
Cerebral Blood Flow in Subjects With Social Phobia During Stressful Speaking Tasks: A PET Study
Abstract
OBJECTIVE: The central nervous system representation of social phobia (social anxiety disorder) is largely unknown. The aim of this study was to examine brain activity during symptom provocation in social phobics. METHOD: Positron emission tomography with the use of 15O water was used to measure regional cerebral blood flow (rCBF) in 18 subjects with DSM-IV-defined social phobia and a nonphobic comparison group while they were speaking in front of an audience and in private. Heart rate and subjective anxiety were also recorded. RESULTS: During public versus private speaking, subjective anxiety increased more in the social phobics than in the comparison group. Increased anxiety was accompanied by enhanced rCBF in the amygdaloid complex in the social phobics relative to the comparison subjects. Cortically, brain blood flow decreased in the social phobics and increased in the comparison subjects more during public than private speaking in the orbitofrontal and insular cortices as well as in the temporal pole and increased less in the social phobics than in the comparison group in the parietal and secondary visual cortices. Furthermore, rCBF increased in the comparison group, but not in the social phobics, in the perirhinal and retrosplenial cortices. CONCLUSIONS: An rCBF pattern of relatively increased cortical rather than subcortical perfusion was observed in the nonphobic subjects, indicating that cortical evaluative processes were taxed by public performance. In contrast, the social phobia symptom profile was associated with increased subcortical activity. Thus, the functional neuroanatomy of social phobia involves the activation of a phylogenetically older danger-recognition system.
A growing body of knowledge links psychopathology to functional brain alterations in anxiety disorders, such as posttraumatic stress disorder (PTSD) (1–3), specific phobia (4–6), panic disorder (7–9), and social phobia (10–12). Through neuroimaging, the orbitofrontal, prefrontal, insular, temporal, cingulate, parietal, and occipital cortices have been identified as important neural network nodes in pathological anxiety. Furthermore, there is a convergence of findings from lesion (13–15), animal (16), and neuroimaging (2, 11, 17–19) studies demonstrating that the amygdaloid complex plays a crucial role in the perception and production of fear. However, the neural correlates of anxiety provocation in patients with social phobia, arguably the most common anxiety disorder (20), have been studied to a limited extent with preliminary data only (10–12). Therefore, the aim of the present study was to examine the functional neuroanatomy of the provocation of social anxiety in subjects with social phobia during a public speaking task, using the same verbal task without an audience for baseline measures. To explore the neural pattern that is specific for social phobia, we included a healthy nonphobic comparison group.
There are several studies of the redistribution of regional cerebral blood flow (rCBF) resulting from perceptually induced anxiety in patients with specific phobia (4, 5, 21, 22), social phobia (11–12), and PTSD (1, 19). We reviewed those studies to establish a priori hypotheses concerning which brain territories to target. During anxiety provocation, increased rCBF has often been observed in the secondary visual cortex (1, 4, 5, 22), except in subjects with social phobia, in whom reductions were observed (12). Because the present study involved anxiety provocation in subjects with social phobia, we predicted involvement of the secondary visual cortex, but we did not predict the direction of change. Enhanced amygdaloid blood flow has been reported in subjects with social phobia during aversive conditions (17, 23) and in normal subjects during fear conditioning (24–26). Since social phobia in part reflects negative learning contingencies related to fear conditioning (27), we predicted increased blood flow in the amygdaloid complex during stressful tasks.
The posterior cingulate cortex, particularly the retrosplenial area, and the inferior frontal cortex have consistently been associated with increased perfusion during normal emotional experiences (28). In individuals with animal phobia, rCBF is down-regulated in the posterior cingulate cortex as a function of anxiety provocation (5, 22), whereas increased posterior cingulate activation has been observed in subjects with social phobia (11) and trauma patients (1). Regional CBF in the anterior cingulate cortex was found to be altered in patients with PTSD (19) and social phobia (11). However, these changes did not have a uniform direction, since both decreased (19) and increased (11) activity have been reported. Thus, we predicted involvement of the posterior and anterior cingulate cortices during symptom provocation but not whether rCBF would increase or decrease.
Frontal activity, including in the prefrontal and orbitofrontal areas, seems attenuated during symptom provocation in most (1, 4, 5, 12, 21, 22) but not all (11, 19) studies of anxiety provocation. A similar pattern is evident for neural alterations in the temporal cortex, since some (1, 4, 5, 22) but not all (11) studies report lower rather than higher rCBF while subjects experience fear. Hence, we predicted lower activity in the frontal and temporal cortices while the subjects were experiencing social anxiety. Finally, we predicted decreased activity in the insular and parietal cortices because the former seems to be involved in a negative feedback loop controlling autonomic nervous system activity while subjects are experiencing aversive conditions (29). Decreased parietal rCBF was also reported by Bremner and colleagues (1) during provocation of posttraumatic anxiety.
In summary, brain areas that have previously been reported to alter neural activity as a consequence of anxiety provocation were investigated in individuals with and without social phobia during public (i.e., speaking in front of an audience) and private (i.e., speaking alone) speaking.
Method
Subjects
Eighteen right-handed individuals (10 men and eight women) with DSM-IV-defined social phobia (mean age=35.2 years, SD=7.3, range=23–46) and six right-handed, healthy, nonphobic subjects (three men and three women) (mean age=22.5 years, SD=1.2, range=21–26) were recruited. The number of individuals with social phobia was higher than the number of nonphobic subjects (N=6). The anxiety-provocation session was repeated after treatment with cognitive behavior therapy (N=6) and citalopram (N=6) as well as for a wait-listed comparison group (N=6) (reported separately), creating four groups of equal size. Criteria for exclusion were previous or current organic brain disorders and somatic diseases, current comorbid axis I disorders, menopause, and pregnancy. The screening procedure included obtaining psychiatric (30) and medical histories. Participants refrained from using tobacco, alcohol, and caffeine 12 hours before the positron emission tomography (PET) scanning. The medical faculty ethical review board and the isotope committee of Uppsala University approved the study. Written informed consent was obtained after the procedure had been fully explained.
Procedure
The subjects lay in the PET scanner with their eyes open during all tasks. During the scanning, the subjects were either speaking in the presence of an audience (consisting of six to eight individuals who were told to remain silent and neutral) or alone for 2.5 minutes. About 20 minutes before the initial scan, the subjects were instructed to prepare a 2.5-min speech about a travel experience or a vacation. Each task was repeated, and the subjects were scanned twice with the order counterbalanced between subjects. During the provocation condition but not during the control condition, the participants were videotaped to enhance their social anxiety. Anxiety was evaluated by measuring their heart rate during each scan, recorded by means of the PSYLAB6 integrated system for psychophysiology (http://www.psylab. com), Spielberger’s State-Trait Anxiety Inventory (state portion only, possible rating=20–80) (31), and ratings of fear and distress (possible rating=0–100) (22) immediately after each scan.
Nervous System Recordings
Subjective and physiological measures of fear and anxiety were averaged over the repetitions for each condition separately and analyzed with repeated measures analyses of variance (ANOVAs) as well as unpaired and paired t tests performed in Statview 4.5 for Macintosh (SAS Institute, Inc.). Because we predicted greater anxiety increases from private to public speaking conditions in the subjects with social phobia than in the comparison subjects, an alpha level of 0.05 was applied.
Blood Flow Recordings
Investigations were performed on a GEMS PC2048-15B scanner (General Electric, Milwaukee, Wis.) with a 10-cm axial field of view (32), producing 15 slices with 6.5 mm of slice spacing and 6 mm of axial/transaxial resolution. A venous catheter was inserted, and the subjects were fixated in the scanner by means of a commercial headholder with fast-hardening foam. First, a 10-min transmission scan with a rotating 68germanium pin source was completed. During the end of the transmission scan, each subject was given a saline injection to attenuate any novelty effects. Before each emission scan, 700–1300 megabecquerels of 15O water in 3–4 ml of water (approximately 15 megabecquerels/kg of body weight) was injected in 12-min intervals. The subjects were told to start speaking immediately after the injections, and data were collected in 15 10-second frames. Each subject had a total of four emission scans during two conditions presented in counterbalanced order between the subjects. Finally, an additional injection was given when the scanner bed was automatically moved back and forth between two positions 10 cm apart. Data were collected only at these positions and never while the scanner was moving. This last interleaved scan was performed to obtain a scan with full axial coverage of the brain, which aids in the stereotactical normalization of PET images.
Data obtained during the first 70 seconds after arrival of the bolus to the brain were summed. Images were reconstructed from the summation after correction for attenuation and scatter by using the transmission scan (33). The additional interleaved scan was reconstructed to a 30-slice image set with 20-cm of coverage in the axial (superior to inferior) direction (34). All images were reconstructed to a 128×128 matrix with a pixel size of 2 mm by using a 15-mm filter.
Normalization of all individual CBF images into a standard brain shape (35) was performed automatically by matching the scan with 20 cm of axial coverage to an atlas template (36). The images from the four emission scans were automatically aligned to the 20-cm scan (37), bringing them into the stereotactic space and correcting for head movements between scans. The stereotactic space was defined on the basis of the postmortem brain slicing of a single subject in whom brain contours, gyri, sulci, central structures, and Brodmann’s areas were defined (35). Software containing a cerebral brain atlas (38) allows for identification, in terms of anatomy, cytoarchitecture, and Talairach coordinates (39).
A group-by-condition blocked ANOVA (40) with one between-group factor (subjects with social phobia versus comparison subjects) and one within-group factor (public versus private speaking) was used. The two normalized rCBF images acquired during each task were averaged into mean images to increase signal-to-noise ratios and to account for between-subject variance, which was compatible with procedures carried out in a random effects model (41). In order to compare rCBF activity from the 18 subjects with social phobia with rCBF activity from the six comparison subjects, we used a balanced contrast vector (40). Global flow was estimated by means of a predefined mask outlining the brain but excluding all voxels that changed as a consequence of study conditions and F-map masking (42). PET data were normalized for global flow by use of linear scaling (42). A threshold z score of 2.6, uncorrected for multiple comparisons (43), was used to examine areas of activation within the brain areas involved in our hypothesis.
Results
Affective Responses
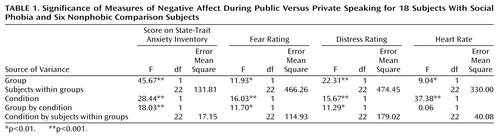
Four repeated ANOVAs (group by condition), including heart rate, State-Trait Anxiety Inventory score (state portion), fear rating, and distress rating as dependent variables, revealed significant main and interaction effects for all the variables except for heart rate (Table 1 and Figure 1). The pairwise planned comparisons revealed significantly higher heart rates during public than private speaking for both subjects with social phobia (t=6.8, df=17, p<0.0001) and nonphobic comparison subjects (t=2.7, df=5, p<0.05). Significantly higher anxiety responses during public than private speaking were exhibited by the subjects with social phobia (State-Trait Anxiety Inventory score: t=8.5, df=17, p<0.0001; fear rating: t=6.6, df=17, p<0.0001; and distress rating: t=6.5, df=17, p<0.0001) but not the nonphobic comparison group (State-Trait Anxiety Inventory score: t=2.1, df=5, p<0.10; fear rating: t=1.5, df=5, n.s.; and distress rating: t=1.7, df=5, n.s.). Furthermore, the subjects with social phobia had higher heart rates (t=2.6, df=22, p<0.05) and anxiety ratings (State-Trait Anxiety Inventory score: t=7.7, df=22, p<0.0001; fear rating: t=3.8, df=22, p<0.001; and distress rating: t=4.8, df=22, p<0.0001) during the public speaking condition than the nonphobic comparison group. During private speaking, the social phobia group displayed significantly higher heart rates (t=3.2, df=22, p<0.005) and anxiety ratings (State-Trait Anxiety Inventory score: t=4.9, df=22, p<0.0001) than the nonphobic comparison subjects, but fear and distress ratings did not differ.
Regional Cerebral Blood Flow
On the basis of our a priori hypotheses, significant local neural alternations with z scores above 2.6 for the interaction contrast with double subtraction (phobicspublic – phobicsprivate) – (comparisonspublic – comparisonsprivate) revealed that rCBF increases were greater in the social phobia group than in the comparison group in the right amygdaloid complex (the amygdala and the periamygdaloid cortex, corresponding to Brodmann’s area 34), extending into the hippocampus (Figure 2 and Table 2). In addition, in the subjects with social phobia, rCBF increases in the right amygdala from public to private speaking were positively correlated with the corresponding change scores for self-reported fear (rxy=0.52, df=16, p<0.05).
Cortically, brain blood flow decreased in the social phobics and increased in the comparison subjects more during public than private speaking bilaterally in the insular cortex as well as in the right temporal pole and increased less in the social phobia group than in the comparison group in the right parietal and secondary visual cortices. Furthermore, rCBF increased in the comparison group, but not in the social phobics, bilaterally in the perirhinal and retrosplenial cortices (Table 2). In addition, rCBF in the orbitofrontal cortex (Brodmann’s area 12) tended to decrease more in subjects with social phobia than in the comparison subjects (maximum z=2.30, N=24; x=3, y=58, z=–9). Plots for the maximum voxel values in the structures displaying significant group-by-condition differences are presented in Figure 3, except for the amygdaloid complex, which is presented in Figure 2.
Discussion
Our aim was to explore the functional neuroanatomy of social phobia by recording rCBF in subjects with social phobia during public and private speaking compared to that in a healthy nonphobic comparison group. The subjects with social phobia spoke in public while experiencing more anxiety than the nonphobic subjects, whereas anxiety differences were smaller during private speaking. Increased fear and anxiety was associated with increased rCBF in the right amygdaloid complex, which is consistent with the findings from previous studies identifying the amygdaloid complex as important for emotion, particularly fear (for a review, see reference 44). The right lateralized activations are in line with theories of emotion that emphasize the right hemisphere as being dominant in experiencing negative affect (45). Furthermore, aversively controlled behavior induces increased long-term potentiation of efferents of the right amygdala in animals (46). Cortically, brain blood flow decreased in the social phobics and increased in the comparison subjects more during public than private speaking in the orbitofrontal and insular cortices as well as in the temporal pole and increased less in the social phobia group than in the comparison group in the parietal and secondary visual cortices. Furthermore, rCBF increased in the comparison group, but not in the social phobics, in the perirhinal and retrosplenial cortices. This neural pattern most likely reflects emotional processes, since the public and private speaking tasks were identical for both groups and were associated with an anxious reaction during public speaking in the subjects with social phobia but not in the nonphobic subjects. Because we evaluated the interaction between the subjects with social phobia and the nonphobic comparison subjects and public versus private performance, rCBF alterations resulting from perceptual or anatomical differences can be ruled out. Instead, they seem to reflect the emotional impact of the situation on the subjects.
Consistent with other PET studies of symptom provocation in subjects with anxiety disorders (2, 11, 19), we observed increased rCBF in the amygdaloid complex during anxiety provocation, which may be consistent with Reiman’s description (11) of a localized alarm center in the amygdala and hippocampus. Hence, social anxiety seems associated with increased activity both in the amygdala and hippocampus. Birbaumer et al. (17) also observed that the amygdala was activated in the processing of neutral faces in subjects with social phobia but not in nonphobic subjects. A complementary explanation for the rCBF activations in the amygdaloid complex could be that evaluation of ambiguous cues (47), such as neutral faces, is supported by the amygdala in subjects with social phobia, whereas cortical areas perform this evaluative process in nonfearful subjects. Thus, there is a fear-related shift from cortical to subcortical processing in subjects with social phobia when encountering situations causing symptom activation.
Individuals with phobic disorders are characterized by an inability to control fear in phobic situations (48). Hence, the rCBF reductions in the orbitofrontal cortex (Brodmann’s area 12), together with enhanced rCBF in the amygdaloid complex, may be associated with emotional dysregulation linked with failure to inhibit negative affect (49). For example, Morgan and co-workers (50) demonstrated that lesions to the medial prefrontal cortex prolonged fear extinction. The orbitofrontal cortex seems particularly involved in voluntary emotional control (5), and lowered orbitofrontal perfusion has also been observed during anxiety provocation in subjects with specific phobia (5, 21, 22), PTSD (51), and panic attacks (52). Because a reciprocal relation between rCBF in the subgenual cingulate and the prefrontal cortex has been described in depression and normal sadness (53), we could speculate that negative affect in general is associated with limbic-cortical reciprocity.
Public speaking anxiety in subjects with social phobia seems associated with altered neural activity in subcortical and cortical structures of relevance for affect and attention. The neural alterations in the present study, located in the secondary visual (Brodmann’s area 19), retrosplenial (Brodmann’s areas 29 and 30), parietal (Brodmann’s area 39), and temporal (Brodmann’s areas 36 and 38) cortices, have previously been suggested to be involved in emotional evaluative processes (4, 11, 28, 54, 55), e.g., visuospatial processing (56). However, perception of novel faces in nonphobic subjects activates a network that includes the right parietal and secondary visual cortices, as well as the right fusiform gyrus (57, 58). Thus, it is likely that the relatively higher neural activity in nonphobic subjects than in subjects with social phobia indicates cortical rather than subcortical evaluative processes. The reverse characterizes subjects with social phobia, and amygdala activation is associated both with perception and generation of emotion (for a review, see reference 44).
Altered rCBF in the insular cortex (Brodmann’s areas 13 and 14) could reflect regulation of emotional autonomic responses (29, 44). For example, we recently reported a negative correlation between insular blood flow and measures of activation in the autonomic nervous system, which suggests a negative feedback function for the insula (29). The pattern of change in the present study also indicates an inverse relation between autonomic anxiety indices and insular perfusion.
In previous PET studies, social anxiety has been associated with 1) enhanced perfusion in the right dorsolateral prefrontal and the left parietal cortices (10), 2) deactivations in the visual and medial frontal cortices (12), and 3) rCBF increases in the lateral prefrontal, sensorimotor, anterior temporal, and midcingulate cortices as well as in the thalamus. Increases that approached significance were reported for the amygdala, hippocampus, hypothalamus, and cerebellar vermis as well as for the anterior cingulate and medial prefrontal cortices (11). Because these studies (10–12) reported only preliminary data and since the completed studies do not seem to have been published, it is difficult to properly evaluate and compare these results with ours.
Traditionally, z scores of 3.09 that are sometimes corrected for multiple comparisons (1) and sometimes not (19) are judged significant. In the present study, areas in which the direction of change was predicted produced z scores that varied from 2.64 to 3.70, whereas areas in which we did not predict the direction of change, only involvement, had z scores exceeding 3.09. A z score of 2.58 corresponds to a one-tailed probability of 0.005 and a score of 3.09 to a two-tailed probability of 0.002. Because rCBF alterations in all areas studied were theoretically predicted on the basis of previous independent research, we believe that they are unlikely to reflect statistical type I errors. Of course, independent replication of the present data would strongly support our findings and interpretations. Although these data are preliminary and were compared with those from a small nonphobic group, the results are consistent with a wealth of data showing that alterations in the amygdala are associated with fear and anxiety (44).
Recently, Paradiso and co-workers (59) reported that observing and assigning emotional value to unpleasant stimuli activated subcortical limbic regions, whereas evaluation of pleasant stimuli activated cortical paralimbic areas. They suggested that the former reflects activity in an archaic danger-recognition system and the latter, a phylogenetically younger system. To conclude, we speculate that subcortical activations observed in the subjects with social phobia during symptom provocation represented anxiety-related activation of this phylogenetically older danger system, whereas cortical activity in the nonphobic subjects represented evaluative activity in a phylogenetically younger system.
 |
 |
Received April 26, 2000; revisions received Sept. 22, 2000, and Jan. 30, 2001; accepted Feb. 12, 2001. From the Department of Psychology and the Uppsala University PET Center, Uppsala University; the Department of Psychiatry, University Hospital, Uppsala, Sweden; and the Psychiatric Neuroimaging Group, Massachusetts General Hospital, Harvard Medical School, Boston. Address reprint requests to Ms. Tillfors, Department of Psychology, Uppsala University, Box 1225, SE-751 42 Uppsala, Sweden, [email protected] (e-mail). Funded by the Swedish Council for Research in the Humanities and Social Sciences and Uppsala University (grants to Dr. Fredrikson) and the Swedish Brain Foundation and the Wenner-Gren Foundation (grants to Dr. Fischer). The authors thank Jesper Andersson and Lennart Thurfjell for discussions and the staff of the Uppsala University PET Center for use of their research site.

Figure 1. Mean Heart Rates and Subjective Measures of Negative Affect During Private Versus Public Speaking for 18 Subjects With Social Phobia and Six Nonphobic Comparison Subjects

Figure 2. Composite PET Scan of Increased Normalized Relative Regional Cerebral Blood Flow (rCBF) in the Amygdaloid Complex During Public Versus Private Speaking for 18 Subjects With Social Phobia and Six Nonphobic Comparison Subjectsa
aLighter colors signify greater differences. Scan is superimposed on a standard magnetic resonance reference image.

Figure 3. Normalized Relative Regional Cerebral Blood Flow (rCBF) in Brain Structures With Alterations During Public and Private Speaking in 18 Subjects With Social Phobia and Six Nonphobic Comparison Subjects
1. Bremner JD, Staib LH, Kaloupek D, Southwick SM, Soufer R, Charney DS: Neural correlates of exposure to traumatic pictures and sound in Vietnam combat veterans with and without posttraumatic stress disorder: a positron emission tomography study. Biol Psychiatry 1999; 45:806-816Crossref, Medline, Google Scholar
2. Rauch SL, van der Kolk BA, Fisler RE, Alpert NM, Orr SP, Savage CR, Fischman AJ, Jenike MA, Pitman RK: A symptom provocation study of posttraumatic stress disorder using positron emission tomography and script-driven imagery. Arch Gen Psychiatry 1996; 53:380-387Crossref, Medline, Google Scholar
3. Shin LM, McNally RJ, Kosslyn SM, Thompson WL, Rauch SL, Alpert NM, Metzger LJ, Lasko NB, Orr SP, Pitman RK: Regional cerebral blood flow during script-driven imagery in childhood sexual abuse-related PTSD: a PET investigation. Am J Psychiatry 1999; 156:575-584Abstract, Google Scholar
4. Fredrikson M, Wik G, Greitz T, Eriksson L, Stone-Elander S, Ericson K, Sedvall G: Regional cerebral blood flow during experimental phobic fear. Psychophysiology 1993; 30:127-131Google Scholar
5. Fredrikson M, Wik G, Annas P, Ericson K, Stone-Elander S: Functional neuroanatomy of visually elicited simple phobic fear: additional data and theoretical analysis. Psychophysiology 1995; 32:43-48Crossref, Medline, Google Scholar
6. Rauch SL, Savage CR, Alpert NM, Miguel EC, Baer L, Breitner HC, Fischman AJ, Manzo PA, Moretti C, Jenike MA: A positron emission tomographic study of simple phobic symptom provocation. Arch Gen Psychiatry 1995; 52:20-28Crossref, Medline, Google Scholar
7. Malizia AL, Cunningham VJ, Nutt DJ: Flumazenil delivery changes in panic disorder at rest (abstract). Neuroimage 1997; 5:302Google Scholar
8. Reiman EM, Raichle ME, Robins E, Mintun MA, Fussleman MJ, Fox PT, Price JL, Hackman KA: Neuroanatomical correlates of a lactate-induced anxiety attack. Arch Gen Psychiatry 1989; 46:493-500Crossref, Medline, Google Scholar
9. Woods SW, Koster K, Krystal JK, Smith EO, Zubal IG, Hoffer PB, Charney DS: Yohimbine alters regional cerebral blood flow in panic disorder (letter). Lancet 1988; 2:678Crossref, Medline, Google Scholar
10. Bell CJ, Malizia AL, Nutt DJ: The neurobiology of social phobia. Eur Arch Psychiatry Clin Neurosci 1999; 249(suppl 1):11-18Google Scholar
11. Reiman EM: The application of positron emission tomography to the study of normal and pathologic emotions. J Clin Psychiatry 1997; 58(suppl 16):4-12Google Scholar
12. Van Ameringen M, Mancini C, Oakman JM, Kamath M, Nahmias C, Szechtman H: A pilot study of PET in social phobia (abstract). Biol Psychiatry 1998; 43(suppl):31SGoogle Scholar
13. Bechara A, Tranel D, Damasio H, Adolphs R, Rockland C, Damasio AR: Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans. Science 1995; 269:1115-1118Google Scholar
14. Broks P, Young AW, Maratos EJ, Coffey PJ, Calder AJ, Isaac CL, Mayes AR, Hodges JR, Montaldi D, Cezayirli E, Roberts N, Hadley D: Face processing impairments after encephalitis: amygdala damage and recognition of fear. Neuropsychologia 1998; 36:59-70Crossref, Medline, Google Scholar
15. Scott SK, Young AW, Calder AJ, Hellawell DJ, Aggleton JP, Johnson M: Impaired auditory recognition of fear and anger following bilateral amygdala lesions. Nature 1997; 385:254-257Crossref, Medline, Google Scholar
16. Cahill L, McGaugh JL: Mechanisms of emotional arousal and lasting declarative memory. Trends Neurosci 1998; 21:273-313Crossref, Medline, Google Scholar
17. Birbaumer N, Grodd W, Diedrich O, Klose U, Erb M, Lotze M, Schneider F, Weiss U, Flor H: fMRI reveals amygdala activation to human faces in social phobics. Neuroreport 1998; 9:1223-1226Google Scholar
18. Breiter HC, Etcoff NL, Whalen PJ, Kennedy WA, Rauch SL, Buckner RL, Strauss MM, Hyman SE, Rosen BR: Response and habituation of the human amygdala during visual processing of facial expression. Neuron 1996; 17:875-887Crossref, Medline, Google Scholar
19. Shin LM, Kosslyn SM, McNally RJ, Alpert NM, Thompson WL, Rauch SL, Macklin ML, Pitman RK: Visual imagery and perception in posttraumatic stress disorder. Arch Gen Psychiatry 1997; 54:233-241Crossref, Medline, Google Scholar
20. Jefferys D: Social phobia: the most common anxiety disorder. Aust Fam Physician 1997; 26:1061, 1064-1067Google Scholar
21. Johanson A, Gustafson L, Passant U, Risberg J, Smith G, Warkentin S, Tucker D: Brain function in spider phobia. Psychiatry Res 1998; 84:101-111Crossref, Medline, Google Scholar
22. Wik G, Fredrikson M, Ericson K, Eriksson L, Stone-Elander S, Greitz T: A functional cerebral response to frightening visual stimulation. Psychiatry Res 1993; 50:15-24Crossref, Medline, Google Scholar
23. Schneider F, Weiss U, Kessler C, Müller-Gärtner H-W, Posse S, Salloum JB, Grodd W, Himmelmann F, Gaebel W, Birbaumer N: Subcortical correlates of differential classical conditioning of aversive emotional reactions in social phobia. Biol Psychiatry 1999; 45:863-871Crossref, Medline, Google Scholar
24. Furmark T, Fischer H, Wik G, Larsson M, Fredrikson M: The amygdala and individual differences in human fear conditioning. Neuroreport 1997; 8:3957-3960Google Scholar
25. LaBar KS, Gatenby JC, Gore JC, LeDoux JE, Phelps EA: Human amygdala activation during conditioned fear acquisition and extinction: a mixed-trial fMRI study. Neuron 1998; 20:937-945Crossref, Medline, Google Scholar
26. Morris JS, Öhman A, Dolan RJ: Conscious and unconscious emotional learning in the human amygdala. Nature 1998; 393:467-470Crossref, Medline, Google Scholar
27. Öst L-G: Ways of acquiring phobias and outcome of behavioral treatments. Behav Res Ther 1985; 23:683-689Crossref, Medline, Google Scholar
28. Maddock RJ: The retrosplenial cortex and emotion: new insights from functional neuroimaging of the human brain. Trends Neurosci 1999; 22:310-316Crossref, Medline, Google Scholar
29. Fredrikson M, Furmark T, Olsson MT, Fischer H, Andersson J, Långström B: Functional neuroanatomical correlates of electrodermal activity: a positron emission tomographic study. Psychophysiology 1998; 35:179-185Crossref, Medline, Google Scholar
30. First MB, Gibbon M, Spitzer RL, Williams JBW: SCID-I and SCID-II, Clinical Version: Interview Protocol in Swedish. Stockholm, Pilgrim Press, 1998Google Scholar
31. Spielberger CD, Gorsuch RL, Lushene R, Vagg PR, Jacobs GA: Manual for the State-Trait Anxiety Inventory. Palo Alto, Calif, Consulting Psychologists Press, 1983Google Scholar
32. Holte S, Eriksson L, Dahlbom M: A preliminary evaluation of the Scanditronix PC2048-15B brain scanner. Eur J Nucl Med 1989; 15:719-721Crossref, Medline, Google Scholar
33. Bergström M, Eriksson L, Bohm C, Blomqvist G, Litton J: Correction for scattered radiation in a ring detector positron camera by integral transformations of the projections. J Comput Assist Tomogr 1983; 7:42-50Crossref, Medline, Google Scholar
34. Bergström M, Litton J, Eriksson L, Bohm C, Blomqvist G: Determination of object contour from projections for attenuation correction in cranial positron emission tomography. J Comput Assist Tomogr 1982; 6:365-372Crossref, Medline, Google Scholar
35. Greitz T, Bohm G, Holte S, Eriksson LA: A computerized brain atlas: construction, anatomical content, and some applications. J Comput Assist Tomogr 1991; 15:26-38Crossref, Medline, Google Scholar
36. Andersson JLR, Thurfjell L: Implementation and validation of a fully autonomic system for intra- and interindividual registration of PET brain scans. J Comput Assist Tomogr 1997; 21:136-144Crossref, Medline, Google Scholar
37. Andersson JLR: A rapid and accurate method to realign PET scans utilizing image edge information. J Nucl Med 1995; 36:657-669Medline, Google Scholar
38. Thurfjell L, Bohm C, Bengtsson E: CBA: an atlas-based software tool used to facilitate the interpretation of neuroimaging data. Comput Methods Prog Biomed 1995; 47:51-71Crossref, Medline, Google Scholar
39. Talairach J, Tournoux P: Co-Planar Stereotaxic Atlas of the Human Brain: Three-Dimensional Proportional System. New York, Thieme Medical, 1988Google Scholar
40. Friston KJ, Holmes AP, Worsley KJ, Poline J-B, Frith CD, Frackowiak RSJ: Statistical parametric maps in functional imaging: a general linear approach. Hum Brain Mapp 1995; 2:189-210Crossref, Google Scholar
41. Petersson KM, Nichols TE, Poline J-B, Holmes AP: Statistical limitations in functional neuroimaging I: non-inferential methods and statistical models. Philos Trans R Soc Lond B Biol Sci 1999; 354:1239-1260Google Scholar
42. Andersson JLR: How to estimate global activity independent of changes in local activity. Neuroimage 1997; 60:237-244Crossref, Google Scholar
43. Kosslyn SM, Shin LM, Thompson WL, McNally RJ, Rauch SL, Pitman RK, Alpert NM: Normal effects of visualizing and perceiving aversive stimuli: a PET investigation. Neuroreport 1996; 7:1569-1576Google Scholar
44. Davidson RJ, Irwin W: The functional neuroanatomy of emotion and affective style. Trends Cogn Sci 1999; 3:11-20Crossref, Medline, Google Scholar
45. Sutton SK, Davidson RJ: Prefrontal brain asymmetry: a biological substrate of the behavioral approach and inhibition systems. Psychol Sci 1997; 8:204-209Crossref, Google Scholar
46. Adamec RE: Transmitter systems involved in neural plasticity underlying increased anxiety and defense: implications for understanding anxiety following traumatic stress. Neurosci Biobehav Rev 1997; 21:755-765Crossref, Medline, Google Scholar
47. Whalen PJ: Fear, vigilance, and ambiguity: initial neuroimaging studies of the human amygdala. Curr Directions Psychol Sci 1998; 7:177-188Crossref, Google Scholar
48. Fredrikson M, Fischer H, Wik G: Cerebral blood flow during anxiety provocation. J Clin Psychiatry 1997; 58(suppl 16):16-21Google Scholar
49. Davey G (ed): Phobias: A Handbook of Theory, Research and Treatment. Chichester, UK, John Wiley & Sons, 1997Google Scholar
50. Morgan MA, Romanski LM, LeDoux JE: Extinction of emotional learning: contribution of medial prefrontal cortex. Neurosci Lett 1993; 163:109-113Crossref, Medline, Google Scholar
51. Bremner JD, Innis RB, Ng CK, Staib SH, Salomon RM, Bronen RA, Duncan J, Southwick SM, Krystal JH, Rich D, Zubal G, Dey H, Soufer R, Charney DS: Positron emission tomography measurement of cerebral metabolic correlates of yohimbine administration in combat-related posttraumatic stress disorder. Arch Gen Psychiatry 1997; 54:246-254Crossref, Medline, Google Scholar
52. Fischer H, Anderson JLR, Furmark T, Fredrikson M: Brain correlates of an unexpected panic attack: a human positron emission tomographic study. Neurosci Lett 1998; 251:137-140Crossref, Medline, Google Scholar
53. Mayberg HS, Liotti M, Brannan SK, McGinnis S, Mahurin RK, Jerabek PA, Silva JA, Tekell JL, Martin CC, Lancaster JL, Fox PT: Reciprocal limbic-cortical function and negative mood: converging PET findings in depression and normal sadness. Am J Psychiatry 1999; 156:675-682Abstract, Google Scholar
54. Heller W, Nitschke JB: The puzzle of regional brain activity in depression and anxiety: the importance of subtypes and comorbidity. Cognition and Emotion 1998; 12:421-447Crossref, Google Scholar
55. Fischer H, Wik G, Fredrikson M: Functional neuroanatomy of robbery re-experience: affective memories studied with PET. Neuroreport 1996; 7:2081-2086Google Scholar
56. Vogt BA, Finch DM, Olson CR: Functional heterogeneity in cingulate cortex: the anterior executive and posterior evaluative regions. Cereb Cortex 1992; 2:435-443Medline, Google Scholar
57. Kim JJ, Andreasen NC, O’Leary DS, Wiser AK, Boles Ponto LL, Watkins GL, Hichwa RD: Direct comparison of the neural substrates of recognition memory for words and faces. Brain 1999; 122:1069-1083Google Scholar
58. Kuskowski MA, Pardo JV: The role of the fusiform gyrus in successful encoding of face stimuli. Neuroimage 1999; 9:599-610Crossref, Medline, Google Scholar
59. Paradiso S, Johnson DL, Andreasen NC, O’Leary DS, Watkins GL, Boles Ponto LL, Hichwa RD: Cerebral blood flow changes associated with attribution of emotional valence to pleasant, unpleasant, and neutral visual stimuli in a PET study of normal subjects. Am J Psychiatry 1999; 156:1618-1629Google Scholar

