
The Interpersonal Dimension of Borderline Personality Disorder: Toward a Neuropeptide Model
Abstract
Abstract
Borderline personality disorder is characterized by affective instability, impulsivity, identity diffusion, and interpersonal dysfunction. Perceived rejection and loss often serve as triggers to impulsive, suicidal, and self-injurious behavior, affective reactivity, and angry outbursts, suggesting that the attachment and affiliative system may be implicated in the disorder. Neuropeptides, including the opioids, oxytocin, and vasopressin, serve a crucial role in the regulation of affiliative behaviors and thus may be altered in borderline personality disorder. While clinical data are limited, the authors propose alternative neuropeptide models of borderline personality disorder and review relevant preclinical research supporting the role of altered neuropeptide function in this disorder in the hope of stimulating more basic research and the development of new treatment approaches.
Borderline personality disorder is a complex disorder associated with substantial morbidity, mortality, and public health costs. Prominent symptoms include suicidal behavior, nonsuicidal self-injury, aggressive outbursts, and emotional reactivity, all of which typically manifest in an interpersonal context (1). For several years, there has been an ongoing discussion about whether impulsive aggression or affective dysregulation is at the core of the disorder. While these factors are important in borderline personality disorder, it is the exquisite interpersonal sensitivity that frequently triggers both dysregulated affect and impulsive behaviors, which suggests that this sensitivity perhaps rests at the core of the disorder and may in turn drive impulsivity and dysregulated affect. Consistent with this conceptualization, Gunderson (2) suggests that disturbances in the interpersonal realm may serve as a potential endophenotype for identifying genetic vulnerabilities in borderline personality disorder, but little research has been done on the neurobiological substrates of the interpersonal sensitivity of the disorder.
It is noteworthy that many symptoms in the interpersonal domain of borderline personality disorder are actually manifestations of intrapersonal difficulties (e.g., difficulty being alone and misperception of the intentions of others as malevolent), and this dimension could perhaps be reconceptualized as "intrapersonal dysfunction." We suggest that an internal feeling of well-being, stability, and self-regulation in borderline personality disorder is tenuous and may rely heavily on a sense of interpersonal contact and connectedness. This vulnerability may be related to an underlying dysregulation of neuropeptides. The critical role of neuropeptides, including the opioids, oxytocin, and vasopressin, in the regulation of affiliative behaviors as well as a sense of well-being suggests that these systems may be implicated in borderline personality disorder and are therefore potentially promising targets for treatment. Indeed, Insel (3) speculates about whether "social attachment is an addictive disorder" in discussing Paul MacLean's notion that substance abuse represents an effort to replace opioids ordinarily provided by social attachments. Given that social attachments are quite problematic in borderline personality disorder, the neuropeptides may be involved. In this article, we present relevant preclinical and clinical data in support of a possible role of neuropeptides in this disorder.
We propose models emphasizing the dysregulation of neuropeptides—in particular, the opioids—in disturbance of homeostatic maintenance of well-being and interpersonal connection. To date, our understanding of the neurobiology of borderline personality disorder is limited and the efficacy of pharmacological interventions for the disorder is disappointing. We hope to stimulate research into the neurobiology of interpersonal sensitivity in the context of borderline personality disorder and to open new avenues of investigation that may provide promise for more effective treatments of this serious disorder.
Phenomenology of Borderline Personality Disorder
Biological and environmental contributions to the development of borderline personality disorder are complex, with no single unifying etiological theory (4, 5). Rather, a number of theoretical models have been proposed, each emphasizing a dimension of the disorder (4, 6, 7–9). Three main symptom clusters or dimensions have been identified (7): 1) behavioral dysregulation, characterized by impulsive aggression (4, 8, 10–16), nonsuicidal self-injury and suicidal behavior (6), substance abuse, recklessness, impulsive spending, and inappropriate displays of anger; 2) affect dysregulation (17–19), characterized by emotional lability, intense negative emotional reactions (e.g., shame), difficulty recovering from disturbing triggers (even seemingly modest ones), and pervasive dysphoria (4, 6, 9, 17, 18); and 3) disturbances in interpersonal relatedness, characterized by turbulent relationships and fear of perceived or real abandonment. Neither behavioral nor affect dysregulation is unique to borderline personality disorder. Behavioral problems are prominent in antisocial personality disorder (15, 16) and intermittent explosive disorder. Affective reactivity (19) and dysphoria occur in axis I mood disorders, particularly bipolar disorder. However, unlike bipolar disorder, the affective experience of chronic emptiness (9) seems to be unique to borderline personality disorder.
The third dimension of borderline symptomatology, disturbance in interpersonal relatedness, serves to differentiate this disorder from axis I and other axis II disorders. Gunderson et al. (20) found that two interpersonal criteria, avoidance of abandonment and unstable relationships, differentiated borderline from other personality disorders. As noted above, symptoms in this domain manifest both intra- and interpersonally, and this too is unique to borderline personality disorder (21).
Neurobiological studies have begun to examine this interaction (21–25). In the intrapersonal domain, cognitive distortions include misperceiving others' intentions, classifying people in extremes as "good" and "bad" (26, 27), misperceiving abandonment threats, attributing malevolent intentions to others where none exist, and inability to understand the mental state of others (i.e., mentalization) (28). Interpersonal manifestations typically include excessive dependency and turbulent relationships. However, these symptoms too can be viewed as manifestations of difficulties in the intrapersonal domain: others may be needed for self-definition and soothing, and thus neediness and hypervigilance to real or perceived abandonment can be readily understood.
Interpersonal Dysfunction as a Core Component of Borderline Personality Disorder
Manifestations of behavioral and affect dysregulation in borderline personality disorder can be viewed in the context of a disrupted sense of self in relation to others. According to Fonagy and Bateman (29), the inability to make sense of self and others is at the core of the disorder and is a result of disrupted attachment in early development, which may depend in part on the developing child's capacities and vulnerabilities as well as environmental influences. This conceptualization is consistent with Gunderson's proposed endophenotype of disturbed relatedness (2, 30). Although common conceptions of the interpersonal difficulties of borderline personality disorder focus on external relationships and DSM-IV defines the interpersonal difficulties of the disorder as "a pattern of unstable and intense interpersonal relationships characterized by alternating between extremes of idealization and devaluation," the symptoms that cluster on this dimension in factor analyses are unstable relationships, identity disturbance, chronic feelings of emptiness, and stress-related paranoid ideation (typically, rejection-related paranoid feelings) (31). All but the first factor relate to internal states and self-regulation and have an impact on relatedness. Identity diffusion can also lead to extreme vulnerability to real or perceived loss of important relationships (21) and overreliance on others to experience a coherent sense of self. This perspective is consistent with an object relations theory of borderline personality disorder (26) as well as Bender and Skodol's conceptualization proposing that lack of self-integration is at the core of the disorder (27).
As previously noted, the interpersonal difficulties of borderline personality disorder appear to be responsible for much of the distress these individuals experience in daily life. Loneliness, perceived rejection, and disruptions in relationships are precipitants to suicide attempts (1, 32), nonsuicidal self-injury, and substance use (33), while the anxious-preoccupied insecure attachment style in borderline personality disorder is associated with longer psychiatric hospitalizations (34).
Healthy individuals develop a stable sense of well-being and self-esteem in the context of nurturing and soothing interpersonal relationships over the course of development. This stable sense of well-being and self-esteem depends on both the interpersonal environment and the innate capacity to internalize these soothing relationships. Because of their exquisite sensitivity to separation and interpersonal vicissitudes, individuals with borderline personality disorder seem to experience their self-esteem as dependent on the availability of important others. When these important figures are unavailable or rejecting, these individuals can experience a plummeting loss of well-being with accompanying feelings of abandonment. The behaviors that result—suicidal expressions, emotional outbursts, and clinginess—appear manipulative and excessive, but to the person with borderline personality disorder they may be survival tactics to maintain self-esteem, given that their sense and integrity of self rely to a large extent on others. It is easier to understand the excessive clinginess, demandingness, and panic at the thought of abandonment in the individual with borderline personality disorder if one considers that preserving interpersonal connectedness serves not only to preserve a key relationship but also, perhaps more importantly, to provide a sense of cohesiveness of self. Indeed, this may be a key to understanding the fear of abandonment, panicky avoidance of being alone, problems with trust in intimate relationships, biases in social judgment, and consequent turbulent relationships and self-injurious behavior in borderline personality disorder.
The opioids have been implicated in feelings of soothing or pleasure as well as in the distress of social separation and exclusion, and oxytocin has been implicated in the establishment of trust and affiliative behaviors. Thus, dysregulation of neuropeptides such as opioids and oxytocin may contribute to deficits in the maintenance of well-being, heightened separation distress, and mistrust in borderline personality disorder. Although we focus here on the interpersonal dimension of borderline personality disorder, dysregulation of neuropeptides may also underlie many of the behavioral and affective symptoms of the disorder.
Models of Interpersonal Dysfunction in Borderline Personality Disorder
The interpersonal dysfunction typically seen in this patient group can be conceptualized within one of several paradigms: 1) as distinct from neurobiology and conceptualized purely in psychological or developmental terms; 2) as an emergent phenomenon stemming from the interpersonal consequences of impulsive aggression and/or affective instability; and 3) as a problem arising from neurobiological vulnerabilities that lead to interpersonal sensitivity, partially distinguishable from those implicated in impulsive aggression and affective instability and manifest most robustly in the interpersonal domain.
In the first model, neurobiological factors are conceptualized as relatively nonspecific and less important, with interpersonal disturbances developing in the context of dysfunctional attachment interactions. Specific stressors include neglect, abuse, and other trauma as well as repeated experiences of feeling invalidated. In such a model, neurobiological factors may be considered in terms of a "black box"—that is, nonspecific mechanisms. We will argue that the neurobiological underpinnings related to the interpersonal dysfunction may be explicable in established neurobiological systems related to affiliation and affect regulation that shape the trajectory of interpersonal development in the context of the specific interpersonal environment.
In the second, interpersonal dysfunction emerges as a consequence of the more fundamental disturbances of affect regulation and impulse dyscontrol (4, 22–25). Thus, individuals with borderline personality disorder are less able to modulate their emotions in the context of interpersonal relationships, so their well-being is experienced as dependent on the availability of important others. They are more likely to become disappointed, feel abandoned, or become enraged in their key relationships. Because they have a low threshold for impulsive behaviors, these emotions are translated into action, including outwardly aggressive or self-destructive behaviors that disrupt these relationships. While these dynamics may indeed contribute to much of the relational turmoil of borderline personality disorder, they do not easily account for the feelings of inner deadness or panicky aloneness these individuals feel when they are separated from the important figures in their life, the repeated self-injurious behaviors when their relationships are disrupted, and their reliance on others to maintain a stable sense of self-esteem.
In the third paradigm, there are specific neurobiological systems (35), including neuropeptides such as the opioids and oxytocin, that are implicated in affiliative and interpersonal behaviors. Dysregulation of these peptides may specifically contribute to the disturbed affiliative behaviors by intensifying the stress of separation, rendering the maintenance of self-esteem and a feeling of well-being more problematic, and diminishing the capacity to trust and respond appropriately to others, thus driving self-destructive behaviors that may provide relief from pain. In conjunction with neurological substrates already implicated in the impulsive aggression and affective dysregulation of borderline personality disorder, these dysregulations may more directly affect the capacity to maintain interpersonal relationships.
Considering these alternative paradigms, we propose a model of borderline personality disorder that incorporates vulnerability or dysregulation of the opioid system. We posit low basal opioid levels in this disorder, with a compensatory supersensitivity of μ-opioid receptors such that transient increases in opioids following painful stimuli lead to heightened responses (21, 36). Low basal opioid levels are reflected clinically in a sense of "inner deadness," chronic dysphoria, and lack of a sense of well-being, all characteristics of borderline personality disorder, while stimulation of the opioid system (e.g., nonsuicidal self-injurious behavior) can result in heightened relief of pain and restoration of a sense of well-being. This model is consistent with observations of reduced β-endorphin and met-enkephalin levels in CSF under basal conditions in individuals with cluster B personality disorders and a history of self-injury (36, 37); increased pain threshold following painful stressors in borderline personality disorder (38, 39); some therapeutic effect of naltrexone, an opioid antagonist, for self-injurious behavior (37); and the clinical observation that individuals with borderline personality disorder tend to overuse opiate pain medications and report a sense of well-being when taking these medications. While opiates alleviate subjective distress associated with reduced endogenous basal opioid levels, naltrexone dampens the reward of self-injurious behavior by blocking opioid receptors. However, this approach does not address the putative baseline deficits in opioids.
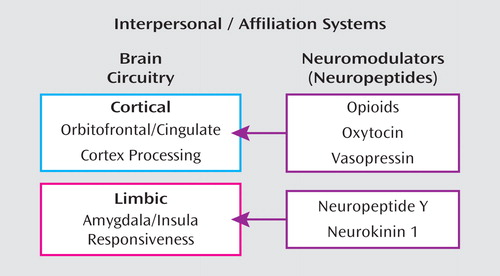
In this model, dysregulation of oxytocin may distort the reading of social cues, appropriate establishment of trust, and capacity for attachment, and vasopressin may in part regulate aggression in the context of intimate relationships. Other neuropeptides outside the purview of this article, such as neuropeptide Y and neurokinin 1, mediate emotional responses to stress mediated by the amygdala, driving affective sensitivity (Figure 1). In this overview, we review relevant preclinical data on neuropeptides such as oxytocin, vasopressin, and opioids, preclinical models of separation distress and social bonding, human studies of these neuropeptides and their associated physiology, clinical studies of self-injurious behavior in borderline personality disorder, and naltrexone trial data. We then elaborate our specific model and conclude by offering suggestions for new research to address the hypotheses and treatment strategies discussed.
The Opioids and Borderline Personality Disorder
Neuropharmacology and Neurophysiology of the Opioid System
The opioid system modulates responses to acute and chronic stressful and noxious stimuli that induce physical, emotional, or social pain. Endogenous opioids include the endorphins, enkephalin, dynorphin, and nociceptin/orphanin FQ agonists. These act on G-protein-coupled receptor subtypes, including μ-opioid receptors, which respond preferentially to morphine; kappa opioid receptors, which respond preferentially to ketocyclazocine; delta receptors; and ORL-1 opioid-like receptor binding sites (40). β-Endorphin and met-enkephalin are agonists at μ-opioid receptors and are related to stress-induced analgesia and thermal pain perception (41–43). Met-enkephalin is also involved in saliency, reward, and motivational behavior (44, 45). μ-Opioid receptors are widely distributed throughout the human CNS, with a particularly dense distribution in the basal ganglia, cortical structures, thalamic nuclei, spinal cord, and specific nuclei in the brainstem (46); high levels of binding of this receptor are found in the basolateral amygdala, nucleus accumbens, hypothalamus, thalamus, ventral tegmental area, and caudate putamen. A variety of stressors activate β-endorphins in rodents (47), and repeated restraint stress reduces opioid receptor density (48). Furthermore, opioid activity activates hypothalamic-pituitary-adrenal (HPA) axis activity, up-regulating corticotropin-releasing factor and pro-opiomelanocortin mRNA (49, 50). μ-Opioid receptors in periaqueductal gray matter mediate the antinociceptive effects of opioids. Thus, the μ-opioid receptor system appears to be particularly relevant for the social and affective regulation associated with borderline personality disorder. The central mediating role of opioids in separation distress, relief and pleasure on reunion, self-soothing, and the pain of social exclusion and rejection suggests that this system is a likely candidate for contributing to the interpersonal vulnerabilities and intrapersonal pain of borderline personality disorder. In the following sections, we review emerging evidence regarding this system in borderline personality disorder.
Opioids, Emotions, and Social Behavior
One hypothesis that provides an explanatory model for the relationship between opioids and emotions posits the mediation of social exclusion, separation, and abandonment, particularly in the context of reactions to perceived rejection, by the same opioid system that modulates physical pain (51). The experience of pain can be divided into painful sensation and painful affect (51, 52). Painful affect appears to be encoded primarily in the anterior cingulate cortex but not in the somatosensory cortex (53). Thus, social exclusion is posited as triggering painful affective feelings without accompanying physical pain.
Brain opioids appear to mediate social affect, particularly social exclusion and separation in the context of social attachments (54). Puppies, young guinea pigs, and chicks emit distress vocalizations in the context of social separation and isolation. These vocalizations can be reduced after administration of opioids, such as oxymorphone. Naloxone, a μ-opioid receptor antagonist, blocks the suppression of distress vocalizations produced by oxymorphone and increases vocalizations several hours later, suggesting a withdrawal effect. Also, administration of low doses of morphine reduces the tendency of animals to spend time close to other members of their species and increases play, especially in socially isolated animals. It is hypothesized that early in the developing brain, opioid systems mediate feelings of soothing and pleasure during early nurturance and later reinforce relief of emotional distress in the context of reunion after separation in meaningful attachments. Low doses of morphine increase play, whereas opioid blockade with naloxone reduces play, which suggests that opioid reduction is associated with reduced pleasure and interaction in vigorous social relationships (55). Interestingly, naloxone has also been found to facilitate increased sexual behavior in some species (56).
With respect to emotional expression, the activation of μ-opioid receptor-mediated neurotransmission has been shown to suppress fear and stress responses to noxious or threatening stimuli and mother-infant separation (57, 58). The μ-opioid receptors also contribute to regulation of emotional memory (59). These animal studies invite comparable human studies to identify more specifically the interpersonal stressors in which opioids play a mediating role.
Human studies have shown that regional activation of μ-opioid neurotransmission is centrally implicated in the suppression of the affective qualities of a pain stressor and in the negative internal affective states induced by that challenge (60, 61). One study demonstrated dynamic changes in μ-opioid neurotransmission in response to an experimentally induced negative affective state (62). The direction and localization of these responses confirm the role of μ-opioid receptors in the regulation of affective experiences in humans. Since the experience of borderline personality disorder is characterized by recurrent stress-induced negative affects, reduced opioid levels may play an important part in enhancing these characteristic negative emotions. It is worth noting that endogenous opioids may mediate the mood-enhancing effect of exercise (60, 61). More specific mapping of the negative affective states modulated by the opioids is indicated in human studies, ideally in an interactional setting.
Opioid Dysfunction and Borderline Personality Disorder
Borderline personality disorder and physical pain.
Affective dysregulation and self-injurious behavior—key characteristics of borderline personality disorder—appear to be associated with lower pain perception. Experimental studies have shown lower pain sensitivity in this patient group (63, 64), as well as significant positive correlations between pain tolerance thresholds, aversive inner tension, and dissociation (65). Psychophysiological research has shown no deficits in the sensory-discriminative aspects of pain processing in borderline personality disorder (38). However, functional imaging research suggests that alterations in activation patterns of brain regions associated with the affective-motivational aspects of pain processing are associated with the pain perception alterations seen in these patients (39). These studies require replication with larger samples.
Borderline symptoms and endogenous opioids.
Endogenous opioids have been implicated in nonsuicidal self-injury, a behavior that occurs frequently in the context of borderline personality disorder (66, 67) and dissociation. Evidence for such a role is based on the partial success of opioid antagonist treatment to ameliorate self-injury (68, 69), as well as on reports of altered pain sensitivity during episodes of self-injurious behavior (70–73) and findings of altered endogenous opioid levels in individuals with histories of self-injury (36, 74, 75). Stanley et al. (36) demonstrated that self-injurers with cluster B personality disorders, predominantly borderline personality disorder, have lower CSF levels of endogenous opioids, in particular β-endorphin and met-enkephalin, compared to those without self-injury. Furthermore, several studies found that naloxone or naltrexone is useful in diminishing nonsuicidal self-injury (76, 77). In a related study focusing on dissociation, another prominent borderline symptom, Bohus et al. (78) found naltrexone to be effective in diminishing dissociation and tonic mobility in borderline personality disorder.
One study suggested that paroxetine, a selective serotonin reuptake inhibitor, induced an antinociceptive effect (79). This antinociception was significantly inhibited by naloxone, suggesting the involvement of opioidergic mechanisms. Specifically, a decrease in negative affect, an increase in positive affect, and an increase in dissociative symptoms were reported. Interestingly, it has been noted in clinical observation that nonsuicidal self-injury in borderline personality disorder is often followed by mood enhancement. Simeon et al. (80) suggested that self-injury could act as self-healing through restoration of positive affect, however brief. This demonstrates the interaction between the serotonergic and opioid systems in the brain. It would be of interest to evaluate the association of self-injury with opioid release using negative affect provocation positron emission tomography imaging paradigms. In summary, multiple lines of evidence suggest that endogenous opioids are involved in the pathogenesis of nonsuicidal self-injury.
μ-Opioid Imaging Studies in Borderline Personality Disorder
Although few studies have been conducted using μ-opioid imaging in borderline personality disorder, the existing investigations point to dysfunction in the disorder. In a study comparing women with borderline personality disorder and healthy comparison women (81), sustained neutral and sadness states were induced by recall of a previously experienced event, and displacement of the μ-opioid receptor selective radiotracer [11C]carfentanil was measured during these states. Women with borderline personality disorder demonstrated greater μ-opioid binding in the orbital frontal cortex, the caudate nucleus, and the nucleus accumbens as well as the left amygdala. During the sadness induction, these women manifested increased opioid release in the right pregenual anterior cingulate, the left orbitofrontal cortex, the left ventral pallidum, the left amygdala, and the left inferior temporal cortex but greater deactivation in the left accumbens, the hypothalamus, and the right hippocampus. These results suggest increased baseline μ-opioid receptor binding and increased sadness-induced opioid release in the orbitofrontal cortex, the caudate, and the accumbens. Paralleling our proposed model of exaggerated response to painful stimuli, increased responsiveness to sadness is consistent with the emotional dysregulation of borderline personality disorder and exaggerated responses to negative experiences or emotions. This is an innovative and important study, but the results must be considered preliminary given the small sample size. Naturalistic interactional paradigms might represent a promising next step in assessing opioid activity in an interpersonal context.
Individual Differences in Opioid Activity
Individual differences in the opioids are grounded in a genetic basis, which includes genetic variation in the opioid receptors as well as opioid release. Against this genetic background, stress, induction of negative affect, and states of acute separation may actively influence opioid concentrations, and developmental differences in rearing may alter the opioid system set-point.
Genetic Studies of the μ-Opioid Receptor, Attachment Behavior, and Borderline Personality Disorder
Recent genetic studies suggest that the μ-opioid receptor gene is associated with attachment abnormalities and borderline personality disorder. Polymorphisms in the μ-opioid receptor gene in humans (OPRM1 AII8G) and rhesus macaques (OPRM, C77G) result in amino acid substitutions in the N-terminal arm of the receptor, leading to increased affinity for β-endorphin in vitro (47, 82) and gain of function in vivo (83, 84), as exemplified in increased alcohol-induced stimulation, an in vivo behavior mediated by endogenous opioids.
In rhesus macaques, the OPRM1 77G allele is associated with higher levels of attachment during early infancy, which suggests increased reward during maternal contact but greater persistence of separation distress (85). The prevalence of this allele is also increased in opioid addicts. Another single-nucleotide polymorphism (SNP) of OPRM1 (rs510769) may be increased in individuals with borderline personality disorder (86) and is associated with affective lability. Interestingly, this SNP accounts for significant variance in prefrontal activation in relation to an aggression provocation task imaged by fluorodeoxyglucose positron emission tomography (L.J. Siever et al., unpublished data). Two SNPs of the opioid delta 1 receptor gene are associated with identity disturbance (86). These data, while quite preliminary, raise the possibility that genetic variability in the opioid receptors may affect affective stability, attachment, and coherence of self-concept. Further investigation of these polymorphisms in larger samples and their relation to functional imaging would help to elucidate these relationships.
Model of Opioid Dysfunction in Borderline Personality Disorder
We propose a model of reduced basal opioid activity in critical limbic circuitry, including the cingulate cortex and the amygdala, in individuals with borderline personality disorder. Observations of reduced CSF concentrations of β-endorphin and met-enkephalin in patients with borderline personality disorder with self-injury, as well as evidence of reduced release in the anterior cingulate cortex, are suggestive of this possibility. μ-Opioid receptors, conversely, are hypothesized to be up-regulated secondary to reduced basal opioids in the amygdala, the striatum, and the orbital frontal cortex, so opioid release would act on supersensitive receptors in these regions. This up-regulation is consistent with regulation of μ-opioid receptors by opioid agonism-antagonism (87, 88). Reduced basal opioids may contribute to the chronic dysphoria and lack of a sense of well-being as well as the difficulties in self-soothing associated with borderline personality disorder. On the other hand, painful stimuli may result in opioid release in the face of more sensitive opioid receptors, thus accounting for observations of a greater pain threshold or a lower pain sensitivity in the disorder. Self-injurious behavior similarly may induce opioid release, stimulating supersensitive opioid receptors and providing a possible mechanism for the sense of well-being and rush often experienced following self-cutting or other forms of self-injury. Drugs that may be abused or prescribed for pain control may enhance basal opioid levels and down-regulate μ-opioid receptors (see Figure 2).
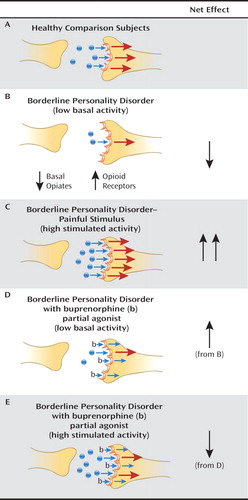
aBasal opioid levels in borderline personality disorder are hypothesized to be reduced in output, while receptors are increased in number, so that during unstimulated conditions, borderline patients experience dysphoria associated with reduced tonic opioid activity. When stress or pain causes an increase in release of opioids, there is an increased opioid signal and relief from dysphoria. Treatment with buprenorphine, a partial agonist, would increase basal opioid signal under baseline conditions of low tonic activity and antagonize opioid receptors under conditions of increased output (e.g., self-injurious behavior).
Lower opioid activity may be a function of genetic factors, environmental factors, or, very likely, a combination of the two. Genetic studies of polymorphism in the opioid receptor suggest abnormalities in eating disorders, which are also associated with impulsive behavior, binge eating, and feelings of dysphoria and satiation. Preliminary data cited earlier suggest that, indeed, there are alterations in opioid receptor genes, including the μ-opioid receptor (OPRM1), associated with affective instability and sensitivity to abandonment (86).
Oxytocin and Borderline Personality Disorder
Neuroanatomy and Neuropharmacology of the Oxytocin System
Oxytocin plays a critical role in maternal behavior, partnering, and a variety of other prosocial behaviors. It is synthesized in magnocellular neurons of the paraventricular and supraoptic nuclei of the hypothalamus (89) and then transported to the posterior pituitary, where it is released (90). Oxytocin receptors are especially abundant in brain areas involved in social behaviors, including the bed nucleus of the stria terminalis, the hypothalamic paraventricular nucleus, the central nucleus of the amygdala, the ventral tegmental area, and the lateral septum (91). In rodent mothers, suckling, as well as visual, auditory, and olfactory cues related to their infants, enhances maternal care behavior in part by increasing expression of oxytocin receptors in these regions (91). In humans, oxytocin is central to affiliative behaviors, including bonding with parents or romantic partners (92, 93). It also plays a key role in emotion regulation and stability. Oxytocin modulates the formation of memories, particularly social and spatial memory, as well as responses to emotional latent stimuli, such as facial expressions. Oxytocin is thus involved in "reading" states in other individuals (i.e., mentalizing) (94). However, there has been limited study of the affiliative role of oxytocin.
Oxytocin and Stress
Oxytocin serves to diminish the stress response (95–97). Lesions of the medial preoptic area eradicate maternal behavioral repertoires. Oxytocin can inhibit HPA axis activity stimulated by stress in women who are lactating (98). Breastfeeding can attenuate the response to stress, suggesting a possible mediating role of oxytocin (98, 99). In general, stimulation of the nipple, as well as social contact, increases oxytocin release and also attenuates HPA axis reactivity (100). In a placebo-controlled double-blind study, healthy men exposed to the Trier Social Stress Test, a laboratory procedure that induces a moderate amount of stress in a social setting, received intranasal oxytocin or placebo before the induced stress, with or without social support from a friend. Both social support and oxytocin diminished the stress response, and the combination of the two resulted in a lower cortisol response (98). Distinct populations of neurons in the amygdala are activated by oxytocin and arginine vasopressin receptor stimulation, and they modulate integration of excitatory data from the amygdala and the cerebral cortex in opposite manners (96).
Oxytocin, Affiliation, and Bonding
Preclinical studies demonstrate oxytocin's crucial role in bonding. Oxytocin receptors are richer in the monogamous prairie voles than in mountain voles, which do not form stable monogamous relationships (101). Oxytocin receptor levels in the central nucleus of the amygdala are significantly higher in rats exhibiting maternal behaviors, including licking, grooming, and nursing of pups, than in those with low levels of these behaviors (91). In humans, oxytocin is a critical mediator of social connectedness and bonding in the context of romantic attachments, enduring monogamous partnerships, and maternal behavior. For example, in romantically unattached young adults, oxytocin is associated with self-report measures of bonding to parents and inversely related to psychological distress (92). Since receptors for oxytocin are more abundant in the reward centers of prairie voles, which form enduring monogamous relationships, it has been hypothesized that oxytocin plays a similar role in adult human romantic attachments (102, 103). While studies of oxytocin and monogamous behavior in humans have not been conducted to date, one study found that in cohabiting couples, greater partner support is associated with higher plasma oxytocin in both men and women before and after a period of warm partner contact (93). In humans, oxytocin concentrations during early pregnancy and during the immediate postpartum period are associated with maternal bonding behaviors, including gaze, vocalizations, positive affect, and affectionate touch, as well as thoughts related to attachment and frequent monitoring of the infant (104). Furthermore, oxytocin mediates attachment behavior over the course of development, with lower urinary concentrations of oxytocin found in maltreated children (105) and in adult males with a history of early separation (106), as well as in the CSF of adult females with a history of childhood abuse (107). However, these few studies are based on retrospective data, such as history of trauma, and the validity of one of the studies (105) has been questioned (108). Therefore, additional studies, particularly with prospective designs, are needed to establish the role of oxytocin in a developmental context.
Oxytocin and Trust
Intranasal administration of oxytocin has been shown in human studies to significantly enhance trust (109, 110). This increase in trust augments social interactions and is based on an increased willingness to accept social risks but not a more generalized increment in readiness to bear risks of all types. This has been demonstrated in a double-blind study design using a trust game in which a participant-investor can transfer money to a trustee and the latter has a choice of sharing the increase generated by the transfer or not. Oxytocin enhances prosocial behavior by influencing the willingness to share in the face of social risks, biasing individuals to increase approach and trust in others (110). One mechanism by which oxytocin may increase prosocial behavior is through influencing affective experiences of others. In a conditioning paradigm, differential negative affective ratings of faces can be induced by aversive conditioning compared to a nonconditioning intervention, and this differential negative evaluation can be reversed by treatment with oxytocin. This effect has been associated with an attenuation of activation in the anterior cingulate cortex and the anterior medial temporal cortex (111). Aversive conditioning increased activation in the extended amygdala, including the anterior medial temporal cortex just anterior to the amygdala, but this effect was not seen in an oxytocin-treated group. Furthermore, activation of the right amygdala is greater for faces with a direct gaze as opposed to an averted gaze in a fear conditioning paradigm, consistent with the social relevance or valence of these faces.
Oxytocin can improve the ability to infer the mental state of others (94), as assessed by a test of inferring the internal state of others from faces with affective facial expressive differences. Oxytocin also improves performance on this task compared with placebo, particularly with more challenging discriminations.
Oxytocin not only influences the recognition of affective cues but also modulates the encoding of positive social interactions into memory. For example, in a double-blind randomized placebo-controlled between-subjects trial (112), healthy males received oxytocin or placebo and then viewed 36 happy, angry, or neutral human faces; they were asked the following day whether they "remembered" or "knew" previously seen faces. Those who received oxytocin were more likely to remember or at least recognize as familiar previously seen happy faces compared with angry and neutral faces, which suggests that oxytocin influences the ongoing representation of positive social interactions.
Oxytocin Dysfunction in Borderline Personality Disorder
Oxytocin is implicated in prosocial behavior, evaluation of others, and ascertainment of others' internal states. To the extent that individuals with borderline personality disorder have difficulty in consciously registering the internal states of others, dysfunction in oxytocin activity may impair their capacity to evaluate others' state of mind from social cues. While limited data are available regarding the biologic activity of oxytocin in borderline personality disorder and its effects, preliminary data suggest that oxytocin reduces stress-induced increases in cortisol in the Trier Social Stress Test (113) and skews responses of patients with borderline personality disorder in a cooperation paradigm (114). In the latter study, borderline patients and healthy comparison subjects played an assurance game emphasizing trust in which the payoff is highest for both players if they cooperate, but if the partner is mistrustful it pays more to defect. In this task, a variant of the prisoner's dilemma game, with a player and a confederate, participants played the assurance game 45 minutes after randomized administration of intranasal oxytocin or placebo. Paradoxically, while comparison subjects tended to cooperate more after administration of oxytocin when the partner was hypothetically cooperative, patients did not; they defected more after receiving oxytocin when paired with the hypothetical cooperative partner. In contrast, they defected less than comparison subjects after receiving oxytocin when the partner is hypothesized to defect. Thus, in healthy comparison subjects, the oxytocin appeared to tune their response to more closely match the speculated intent of their partner, while in the patients with borderline personality disorder, oxytocin seemed to do the converse. These data could be interpreted in the following manner. Individuals with borderline personality disorder may view defection by their partner as a threat, so they appeal to the partner by cooperating, while partner cooperation may signal that there is no threat, offering them the opportunity to gain points for themselves. This interpretation suggests that individuals with borderline personality disorder may view relationships as competitive struggles rather than as collaborative ventures. However, this is a preliminary study with a modest sample size, so further research is needed.
In a laboratory study using a social exchange game in which cooperation benefits two individuals playing the game, King-Casas et al. (115) found that individuals with borderline personality disorder experienced difficulty maintaining cooperation and repairing relationships after cooperation was broken. Neurally, activity in the anterior insula, a region known to respond to norm violations, distinguished healthy participants from those with borderline personality disorder, with healthy individuals showing a strong linear relationship between anterior insula response and both magnitude of monetary offer received from their partner (input) and the amount of money repaid to their partner (output). In contrast, activity in the anterior insula of individuals with borderline personality disorder was related only to the magnitude of payment and not to offers of payment.
Given the specific role of oxytocin in trust and in reading others' internal states—precisely qualities that may be distorted in people with borderline personality disorder—further investigations into this neuropeptide system using games that involve separation are warranted. In previous studies, participants could not view the facial expressions of their "partners" in computerized exercises, so the interpersonal context was not accurately replicated. Innovative strategies involving two partners and using real-time affective interactions and observation of effects would further elucidate the role of oxytocin in borderline personality disorder.
Individual Differences in Oxytocin Activity
Levels of both oxytocin release and oxytocin receptors are genetically regulated, and several alleles for the oxytocin receptor have been identified. Early stress can interfere with the developing neuropeptide system and alter oxytocin receptor binding (103), also modulating its activity.
Oxytocin, Genetics, and Borderline Personality Disorder
Several polymorphisms have been identified in oxytocin genes and have been associated variously with eating disorders and other psychopathology. However, little research on oxytocin genes has been conducted in relation to borderline personality disorder. In the only analysis to date on personality disorders, one of four SNPs for oxytocin tasks (rs877172) was significantly associated with inappropriate intense anger in individuals with personality disorders (86). These preliminary results raise the possibility that there may be genetic differences in oxytocin expression in such patients, but this remains to be definitively demonstrated.
The Vasopressin System and Borderline Personality Disorder
Neuroanatomy and Neuropharmacology of the Vasopressin System
Arginine vasopressin, which is also implicated in social behavior, is synthesized in the hypothalamus supraoptic nucleus and paraventricular nuclei in magnocellular cells whose axons then extend to the posterior pituitary, from which the vasopressin is released into the bloodstream when appropriately stimulated. Parvocellular neurons also may contain vasopressin. The arginine vasopressin gene is on chromosome 20 and is evolutionarily conserved. There are three major receptor subtypes for vasopressin: the AVPR1A, AVPR1B, and AVPR2 receptors (116). AVPR1A transcripts are found widely in the nervous system, including a variety of limbic and subcortical structures.
Vasopressin and Social Behavior
The AVPR1A receptor plays a major role in the modulation of social behavior within the vasopressin system; it determines monogamous partner preference, nurturance of offspring, and selective aggression toward male competitors (117). In mammals, vasopressin is more involved in male behaviors than in female behaviors, which are modulated through oxytocin (117).
Vasopressin also promotes behaviors that are adaptive for monogamous relationships, including paternal care of offspring, protection of mate, and selective preference for a mate in male voles, but vasopressin does not have this role in nonmonogamous species. Vasopressin has a circadian rhythm of release, with peak release during the day, and it appears to have an inhibitory role in corticotropin-releasing hormone and ACTH secretion (118).
Vasopressin also plays a critical role in aggression, primarily aggression between males in monogamous species, such as prairie voles. Vasopressin may be particularly critical in regulating the aggression in the context of peer bonding, so that higher levels of aggression are displayed toward unfamiliar males of the species. Increased interactions with fathers of newborn mice are associated with higher levels of aggression than mice receiving less paternal interaction (119).
Increasing vasopressin 1A receptors in nonmonogamous vole species facilitates the formation of selective expression for a mate, which suggests that this receptor plays an important role in modulating social behavior (120). Indeed, the vasopressin 1A receptor gene is highly polymorphic, such that variability in microsatellite-like elements that control expression differences are associated with differences in social behavior in the laboratory setting (121). Increased binding of this receptor may be enhanced by repeated aggressive encounters in dominant hamsters (122), depending on the species. Agonists and antagonists of this receptor can modulate the expression of aggression (116). Treatment with fluoxetine diminishes offensive aggression in part by blocking the activity of the arginine vasopressin system, which suggests that serotonin antagonizes the aggression-enhancing effects of vasopressin (123). Vasopressin infusions enhance pair bonding by facilitating partner preferences (124). Antagonists inhibit such preferences. This facilitation, however, does not occur in nonmonogamous species of the vole. Increasing vasopressin can induce paternal behavior in the nonmonogamous and aggressive meadow vole (125).
Finally, like oxytocin, vasopressin plays a role in social recognition, as vasopressin 1A and 1B receptor knockout mice may have impaired social recognition (126) and antagonists of vasopressin in normal rats impair social recognition (116).
Vasopressin and Aggressive Behavior
In contrast to the wealth of preclinical data regarding vasopressin, there is a dearth of human studies. CSF vasopressin concentration is positively correlated with a history of disinhibited aggression, including temper tantrums and physical aggression in patients with intermittent explosive disorder, many of whom have comorbid borderline personality disorder (127). However, one study found no differences in CSF vasopressin between violent offenders and comparison subjects (128), which raises the possibility that the vasopressin increases may be particularly associated with aggression in an interpersonal context in those who are interpersonally sensitive and not in antisocial individuals lacking interpersonal sensitivity. Reduced concentrations of antibodies to vasopressin are more common in individuals with conduct disorder (129), which also supports the relationship between vasopressin and interpersonal aggression. Further studies show that administration of intranasal vasopressin increases the perception of threat in response to neutral stimuli (130), which is consistent with the tendency of patients with borderline personality disorder to interpret neutral faces as potentially threatening (131), while elevations in vasopressin have been associated with depression and anxiety (116).
Individual Differences in Vasopressin Activity
Genetic polymorphisms have been identified in AVPR1a and have been found to modulate human social behavior and physiology, specifically prosocial or altruistic decision making (132, 133), prepulse inhibition (134), pair bonding (135), and onset of sexual behavior (136). In humans, developmental influences on vasopressin and its receptor have not been clearly documented, but early social experience (137), manipulations of oxytocin (138), and early maternal separation (139) appear to modulate vasopressin through epigenetic mechanisms, gene expression, and regulation of peptide release.
Model of Oxytocin and Vasopressin Activity in Borderline Personality Disorder
Since oxytocin is involved in affiliation and trust, it would be tempting to propose a simplistic model of oxytocin deficiency in borderline personality disorder. While oxytocin did appear to reduce stress in a pilot study of the Trier Social Stress Test, its effects on a cooperation paradigm are more complex, which suggests that oxytocin actually makes patients with borderline personality disorder hypersensitive to social stimuli, such that increased tuning in to others' motivations is actually aversive. This possibility needs to be tested across a variety of laboratory paradigms.
Vasopressin in CSF is correlated with aggression in individuals with personality disorders, while vasopressin concentrations are inversely associated with prolactin responses to fenfluramine, an index of serotonergic capacity (127), which suggests that these two systems may interact reciprocally to modulate aggression. Individuals with borderline personality disorder who have reduced serotonergic activity may have increased vasopressin concentrations associated with aggression toward peers, consistent with their lowered threshold for anger and aggression. Vasopressin thus may mediate the enhanced irritability and aggression of these individuals in the context of close interpersonal relationships.
Implications for Research and Treatment
Research Implications
Despite the potential promise of understanding the neurobiological basis of the interpersonal sensitivities and vulnerability in borderline personality disorder, there has been surprisingly little investigation in this area. There is still a dearth of psychopharmacologic and psychosocial treatment options with high efficacy for this disorder. A better understanding of the substrates of the relational instability of borderline personality disorder might enhance our treatment options. Neuromodulators such as the neuropeptides that have been shown to be involved in affiliative and relational behaviors could be studied in naturalistic paradigms in interpersonal settings and in response to specific behavioral tasks that involve perception of trust and cooperation versus conflict. In addition to documenting the role of neuropeptides such as oxytocin or opioids, the effects of the administration of neuropeptides such as oxytocin and buprenorphine can also be observed in both naturalistic and laboratory behavioral paradigms. Oxytocin and synthetic oxytocin endogens, such as carbetocin (140, 141), have been administered for labor induction and postpartum hemorrhage and can be administered in the context of paradigms involving affiliation and trust. Vasopressin and its synthetic analogue terlipressin (142) have been administered for septic shock and could be used for laboratory tasks and behavioral imaging paradigms involving elicitation of aggression. Buprenorphine, an opioid agonist, is a candidate for naturalistic provocation paradigms involving interpersonal interaction, self-regulation, and affective regulation. Neuroimaging studies can evaluate regional activation patterns related to interpersonal vicissitudes and affiliative behaviors, especially in the context of paradigms using neuropeptide administration or naturalistic measures in neuropeptide activity. In these ways, individual differences in neuropeptide activity and effect on relevant brain circuitry can be documented.
Naturalistic studies involving measurements of neuropeptides as well as administration of neuropeptides and their analogues are particularly important because they may help in the development of a more specific understanding of the mechanisms of these neuropeptides. For example, studies of opioid release in relation to negative affect and stress implicate these systems in the dysregulation of negative affect in borderline personality disorder (81). Results of oxytocin administration in the context of a trust paradigm in borderline personality disorder suggest that a simplistic hypothesis of reduced oxytocin may not capture the complexity of the influence of oxytocin on affiliative systems and trust in this disorder (114). While vasopressin has been implicated in aggression in personality disorders, there have been no naturalistic studies of vasopressin administration in borderline personality disorder. Thus, at this point, the need for more studies exploring these mechanisms is evident. Innovative studies incorporating imaging and, if possible, a variety of behavioral paradigms involving cooperation, trust, and affective regulation are indicated.
Neuroimaging studies can evaluate components of neuropeptide systems by measuring neuropeptide release or receptors through displacement paradigms and receptor binding studies. Functional neuroimaging studies can also evaluate patterns of regional activation with a focus on relevant limbic regions, such as the anterior cingulate gyrus and the amygdala, in relation to affect regulation, cooperation, and affiliative behaviors using behavioral laboratory paradigms with and without administration of specific neuropeptides. In this way, the relevant circuitry of the neuropeptides could be mapped and their functional implications evaluated.
As discussed, little information is available about individual differences in neuropeptide activity and the effects of neuropeptides on the relevant brain circuitry. Longer-term neuropeptide administration might be used in psychopharmacologic treatment studies (see below). The study of these neuropeptides in the psychiatric disorders is only in its infancy, and little is known about the association of any of them with psychiatric disorders. Thus, the epidemiology and the predictive value of neuropeptide activity is virtually unknown in psychiatric populations. As newer and better paradigms evolve to evaluate neuropeptide activity in psychiatric disorders, it may be possible to evaluate the extent and specificity of associations with any psychiatric disorder.
It is important to note that many biological correlates of psychiatric disorders are actually consequences of the disorders, not causes. This may be the case with the neuropeptides. Rutter (143) elegantly described the need to test the causality of "risk factors" for psychiatric disorders and identified several paradigms for determining causality. Among these paradigms are natural experiments, such as twin and migration studies, to disentangle genetic and environmental effects; avoidance of selection bias designs in which all individuals receive the same "treatment"; and mediational studies that examine differential effects of mediation on subgroups. Ultimately, causality tests should be applied to the study of neuropeptides in borderline personality disorder. It will also be crucial to distinguish between what neuropeptide abnormalities are a consequence of early childhood trauma, which has a high reported incidence in borderline personality disorder, and which disturbances are more directly attributable to the disorder.
It is also noteworthy that the findings on neuropeptides must ultimately be placed in the context of other known risk processes for borderline personality disorder. For example, research is needed to compare the strength of any neuropeptide associations with borderline personality disorder with the strength of associations with other known risk factors. Further research should also determine whether the effects of neuropeptides on borderline personality disorder are related to or depend on other risk factors. This type of information will help to determine whether it is worthwhile to pursue a neuropeptide-based treatment approach.
Treatment Implications
Effects of treatment, both psychosocial and psychopharmacologic, can be observed in naturally occurring or provoked variations in the neuropeptides of interest as well as in response to exogenous neuropeptide administration. A variety of psychosocial treatment options have demonstrated some efficacy in borderline personality disorder–dialectical behavioral therapy (144), cognitive therapy, transference-based therapy (145), mentalization therapy (146), and schema-focused therapy (147, 148)—although these treatments have typically shown efficacy for single symptoms. For example, dialectical behavior therapy reduces suicidal behavior and nonsuicidal self-injury, and transference-focused therapy shows efficacy for reflective functioning but not suicidal behavior or self-injury. Psychopharmacologic interventions may be useful in targeting dimensional vulnerabilities, including impulsive aggression, affective lability, and potentially even the interpersonal turmoil of borderline personality disorder, and thus they may have a broad-based impact on the disorder. Successful treatments may modulate or normalize effects of these neuropeptides on behavior and subjective state so that provocation studies could be done before and after treatment. Agents that modulate these systems may also be used for longer-term treatment. For example, buprenorphine, an opioid partial agonist, is a promising candidate and might be used to enhance basal opioid activity, yet because of its partial agonist properties, it may also serve to block increased endogenous opioid activity. Intranasal oxytocin may serve to enhance trust in certain paradigms, although, as preliminary data suggest, in some contexts it might serve to enhance social sensitivity to the point where it leads to aversive behaviors. Vasopressin has been implicated in aggression, affiliation, pair bonding, anxiety regulation, and social recognition (116, 149, 150). Vasopressin (V1A receptor) antagonism can reduce amygdala activation (151). Vasopressin antagonists have been considered for treatment of major depressive disorder (152) and may be helpful in borderline personality disorder. Clearly, further research is indicated to clarify optimal strategies either to manipulate neuropeptide concentrations or to use them as potential markers of beneficial treatment effects.
Ultimately, a better understanding of these mechanisms will enhance our appreciation of the dynamics of the interpersonal vulnerabilities of individuals with borderline personality disorder by more accurately pinpointing the precise nature of these vulnerabilities and providing a heuristic framework from which to explore the exquisite sensitivity to interpersonal vicissitudes and the related difficulties in maintaining a homeostatic sense of well-being in this disorder.
Conclusions
The evidence reviewed here supports the testing of our proposed model of altered neuropeptide function underlying the interpersonal susceptibilities of borderline personality disorder. Areas of investigation can include 1) examination of basal deficits and receptor supersensitivity of the opioid system contributing to the chronic dysphoria, feelings of emptiness, and sensitivity to rejection and abandonment of borderline personality disorder and the relief from these feelings following self-injurious behaviors or administration of opioids; 2) dysregulation of oxytocin contributing to the mistrustful and antagonistic interpersonal modes associated with borderline personality disorder; and 3) increases in vasopressin associated with irritability and aggression, particularly in the context of close interpersonal relationships. The opioids, vasopressin, and oxytocin merit further investigation for their role in borderline personality disorder and are candidates for possible treatments. Exploring new avenues for treatment is particularly important for this disorder because current approaches are far from satisfactory. While treatment with these agents is premature and fraught with clinical complexities, preliminary research interventions in the opioid systems with agents such as buprenorphine or those targeting the oxytocin or vasopressin systems could be explored to identify better treatments, genetic and epigenetic factors, biomarkers, and early stressors that may presage the interpersonal susceptibilities of borderline personality disorder, with implications for early interventions.
1 : Interpersonal precipitants and suicide attempts in borderline personality disorder. Suicide Life Threat Behav 2006; 36:313–322 Crossref, Medline, Google Scholar
2 : Disturbed relationships as a phenotype for borderline personality disorder (commentary). Am J Psychiatry 2007; 164:1637–1640 Link, Google Scholar
3 : Is social attachment an addictive disorder? Physiol Behav 2003; 79:351–357 Crossref, Medline, Google Scholar
4 : A psychobiological perspective on the personality disorders. Am J Psychiatry 1991; 148:1647–1658 Link, Google Scholar
5 : Suicidal and self-injurious behavior in borderline personality disorder: a self-regulation model, in Understanding and Treating Borderline Personality Disorder: A Guide for Professionals and Families. Edited by GundersonJG HoffmanPD . Washington, DC, American Psychiatric Publishing, 2005, pp 43–63 Google Scholar
6 : The essential nature of borderline psychopathology. J Pers Disord 2007; 21:518–535 Crossref, Medline, Google Scholar
7 : Confirmatory factor analysis of DSM-IV criteria for borderline personality disorder: findings from the Collaborative Longitudinal Personality Disorders Study. Am J Psychiatry 2002; 159:284–290 Link, Google Scholar
8 : Neurobiology of aggression and violence. Am J Psychiatry 2008; 165:429–442 Link, Google Scholar
9 : Are the interpersonal and identity disturbances in the borderline personality disorder criteria linked to the traits of affective instability and impulsivity? J Pers Disord 2001; 15:358–370 Crossref, Medline, Google Scholar
10 : Blunted prefrontal cortical fluorodeoxyglucose positron emission tomography response to meta-chlorophenylpiperazine in impulsive aggression. Arch Gen Psychiatry 2002; 59:621–629 Crossref, Medline, Google Scholar
11 : Serotonin function in human subjects: intercorrelations among central 5-HT indices and aggressiveness. Psychiatry Res 1997; 73:1–14 Crossref, Medline, Google Scholar
12 : Serotonergic studies in patients with affective and personality disorders: correlates with suicidal and impulsive aggressive behavior. Arch Gen Psychiatry 1989; 46:587–599 Crossref, Medline, Google Scholar
13 : Impulsive aggression in borderline personality disorder. Curr Psychiatry Rep 2000; 2:56–61 Crossref, Medline, Google Scholar
14 : Fenfluramine challenge in unipolar depression with and without anger attacks. Psychiatry Res 2000; 94:9–18 Crossref, Medline, Google Scholar
15 : Linking antisocial behavior, substance use, and personality: an integrative quantitative model of the adult externalizing spectrum. J Abnorm Psychol 2007; 116:645–666 Crossref, Medline, Google Scholar
16 : Control of human aggression: a comparative perspective. Am Psychol 1993; 48:16–25 Crossref, Medline, Google Scholar
17 : Emotion dysregulation and the development of borderline personality disorder. Dev Psychopathol 2005; 17:899–925 Crossref, Medline, Google Scholar
18 : Dialectical behavior therapy for treatment of borderline personality disorder: implications for the treatment of substance abuse. NIDA Res Monogr 1993; 137:201–216 Medline, Google Scholar
19 : The nature of borderline personality disorder: multiple dimensions, multiple symptoms, but one category. J Pers Disord 2007; 21:457–473 Crossref, Medline, Google Scholar
20 : Borderline personality disorder, in DSM-IV Sourcebook, vol 2. Edited by WidigerTA FrancesAJ PincusHA RossR FirstMB DavisWW . Washington, DC, American Psychiatric Association, 1996, pp 717–733 Google Scholar
21 : The neurobiology of personality disorders: implications for psychoanalysis. J Am Psychoanal Assoc 2009; 57:361–398 Crossref, Medline, Google Scholar
22 : Neurotransmitter dysfunction in patients with borderline personality disorder. Psychiatr Clin North Am 2000; 23:27–40 Crossref, Medline, Google Scholar
23 : Evidence of abnormal amygdala functioning in borderline personality disorder: a functional MRI study. Biol Psychiatry 2001; 50:292–298 Crossref, Medline, Google Scholar
24 : Amygdala hyperreactivity in borderline personality disorder: implications for emotional dysregulation. Biol Psychiatry 2003; 54:1284–1293 Crossref, Medline, Google Scholar
25 : Neural correlates of emotion processing in borderline personality disorder. Psychiatry Res 2009; 172:192–199 Crossref, Medline, Google Scholar
26 : The Personality Disorders Institute/Borderline Personality Disorder Research Foundation randomized control trial for borderline personality disorder: rationale, methods, and patient charactersitics. J Pers Disord 2004; 18:52–72 Crossref, Medline, Google Scholar
27 : Borderline personality as a self-other representational disturbance. J Pers Disord 2007; 21:500–517 Crossref, Medline, Google Scholar
28 : Mentalizing and borderline personality disorder. J Ment Health 2007; 16:83–101 Crossref, Google Scholar
29 : The development of borderline personality disorder: a mentalizing model. J Pers Disord 2008; 22:4–21 Crossref, Medline, Google Scholar
30 : BPD's interpersonal hypersensitivity phenotype: a gene-environment-developmental model. J Pers Disord 2008; 22:22–41 Crossref, Medline, Google Scholar
31 American Psychiatric Association: Diagnostic and Statistical Manual of Mental Disorders, 4th ed (DSM-IV). Washington, DC, American Psychiatric Association, 1994 Google Scholar
32 : Aversive tension in patients with borderline personality disorder: a computer-based controlled field study. Acta Psychiatr Scand 2005; 111:372–379 Crossref, Medline, Google Scholar
33 : High-risk situations associated with parasuicide and drug use in borderline personality disorder. J Pers Disord 2002; 16:561–569 Crossref, Medline, Google Scholar
34 : Attachment dimensions as predictors of medical hospitalizations in individuals with DSM-IV cluster B personality disorders. J Pers Disord 2004; 18:595–603 Crossref, Medline, Google Scholar
35 : Recent advances in the biological study of personality disorders. Psychiatr Clin North Am 2008; 31:441–461 Crossref, Medline, Google Scholar
36 : Nonsuicidal self-injurious behavior, endogenous opioids, and monoamine neurotransmitters. J Affect Disord (in press) Google Scholar
37 : Biological models of non-suicidal self-injury, in Understanding Nonsuicidal Self-Injury: Origins, Assessment, and Treatment. Edited by NockMK . Washington, DC, American Psychological Association, 2009, pp 99–116 Crossref, Google Scholar
38 : Differential nociceptive deficits in patients with borderline personality disorder and self-injurious behavior: laser-evoked potentials, spatial discrimination of noxious stimuli, and pain ratings. Pain 2004; 110:470–479 Crossref, Medline, Google Scholar
39 : Neural correlates of antinociception in borderline personality disorder. Arch Gen Psychiatry 2006; 63:659–667 Crossref, Medline, Google Scholar
40 : Peripheral analgesic actions of opioids. J Pain Symptom Manage 1991; 6:119–124 Crossref, Medline, Google Scholar
41 : Involvement of spinal opioid systems in footshock-induced analgesia: antagonism by naloxone is possible only before induction of analgesia. Brain Res 1982; 242:309–326 Crossref, Medline, Google Scholar
42 : Endogenous opioids: biology and function. Annu Rev Neurosci 1984; 7:223–255 Crossref, Medline, Google Scholar
43 : Spinal kappa-opioid receptor-mediated antinociception is stimulus specific. Eur J Pharmacol 1987; 137:197–205 Crossref, Medline, Google Scholar
44 : Molecular cloning and functional expression of a mu-opioid receptor from rat brain. Mol Pharmacol 1993; 44:8–12 Medline, Google Scholar
45 : Role of dynorphin and enkephalin in the regulation of striatal output pathways and behavior. Exp Brain Res 1998; 123:60–76 Crossref, Medline, Google Scholar
46 : Subtraction autoradiography of opiate receptor subtypes in human brain. Brain Res 1987; 418:343–348 Crossref, Medline, Google Scholar
47 : Beta-endorphin immunoreactivity in rat plasma: variations in response to different physical stimuli. Life Sci 1981; 29:1669–1674 Crossref, Medline, Google Scholar
48 : Repeated restraint stress reduces opioid receptor binding in different rat CNS structures. Neurochem Res 2005; 30:1–7 Crossref, Medline, Google Scholar
49 : Reorganization of dorsal root ganglion neurons following chronic sciatic nerve constriction injury: correlation with morphine and lidocaine analgesia. Eur J Pharmacol 2007; 568:124–133 Crossref, Medline, Google Scholar
50 : The nociceptin/orphanin FQ antagonist UFP-101 differentially modulates the glucocorticoid response to restraint stress in rats during the peak and nadir phases of the hypothalamo-pituitary-adrenal axis circadian rhythm. Neuroscience 2007; 147:757–764 Crossref, Medline, Google Scholar
51 : Why does social exclusion hurt? the relationship between social and physical pain. Psychol Bull 2005; 131:202–223 Crossref, Medline, Google Scholar
52 : The Challenge of Pain, 3rd ed. London, Penguin, 1996 Google Scholar
53 : Pain affect encoded in human anterior cingulate but not somatosensory cortex. Science 1997; 277:968–971 Crossref, Medline, Google Scholar
54 : The biology of social attachments: opiates alleviate separation distress. Biol Psychiatry 1978; 13:607–618 Medline, Google Scholar
55 : Endogenous opioids and social behavior. Neurosci Biobehav Rev 1980; 4:473–487 Crossref, Medline, Google Scholar
56 : Inhibitory role of opioid peptides in the regulation of aggressive and sexual behaviors in male Japanese quails. J Exp Zool 1997; 277:146–154 Crossref, Medline, Google Scholar
57 : Opiate modulation of separation-induced distress in non-human primates. Brain Res 1988; 440:285–292 Crossref, Medline, Google Scholar
58 : Behavioral and physiologic effects of CRH administered to infant primates undergoing maternal separation. Neuropsychopharmacology 1989; 2:97–104 Crossref, Medline, Google Scholar
59 : Norepinephrine release in the amygdala in response to footshock and opioid peptidergic drugs. Brain Res 1998; 808:134–140 Crossref, Medline, Google Scholar
60 : Mu-opioid receptor-mediated antinociceptive responses differ in men and women. J Neurosci 2002; 22:5100–5107 Crossref, Medline, Google Scholar
61 : Regulation of human affective responses by anterior cingulate and limbic mu-opioid neurotransmission. Arch Gen Psychiatry 2003; 60:1145–1153 Crossref, Medline, Google Scholar
62 : COMT val158met genotype affects mu-opioid neurotransmitter responses to a pain stressor. Science 2003; 299:1240–1243 Crossref, Medline, Google Scholar
63 : Pain perception in self-injurious borderline patients: naloxone effects. Biol Psychiatry 1994; 35:207–209 Crossref, Medline, Google Scholar
64 : Pain perception during self-reported distress and calmness in patients with borderline personality disorder and self-mutilating behavior. Psychiatry Res 2000; 95:251–260 Crossref, Medline, Google Scholar
65 : Elevated pain thresholds correlate with dissociation and aversive arousal in patients with borderline personality disorder. Psychiatry Res 2004; 149:291–296 Crossref, Google Scholar
66 : Neurobiological correlates of borderline personality disorder. Psychopharmacol Bull 2002; 36:69–87 Medline, Google Scholar
67 : The physiology and neurochemistry of self-injurious behavior: a nonhuman primate model. Front Biosci 2005; 10:1–11 Crossref, Medline, Google Scholar
68 : Naltrexone attenuates self-injurious behavior in mentally retarded subjects. Biol Psychiatry 1990; 27:741–746 Crossref, Medline, Google Scholar
69 : Naltrexone and self-injurious behavior: a retrospective population study. J Clin Psychopharmacol 1996; 16:389–394 Crossref, Medline, Google Scholar
70 : Self-injurious behavior and mood regulation in borderline patients. J Pers Disord 1997; 11:146–157 Crossref, Medline, Google Scholar
71 : EEG theta activity and pain insensitivity in self-injurious borderline patients. Psychiatry Res 1999; 89:201–214 Crossref, Medline, Google Scholar
72 : Pain perception in self-injurious patients with borderline personality disorder. Biol Psychiatry 1992; 32:501–511 Crossref, Medline, Google Scholar
73 : Subtypes of self-injurious patients with borderline personality disorder. Am J Psychiatry 1993; 150:1869–1871 Link, Google Scholar
74 : Raised plasma metenkephalin in patients who habitually mutilate themselves. Lancet 1983; 2:545–546 Crossref, Medline, Google Scholar
75 : Dissociation of POMC peptides after self-injury predicts responses to centrally acting opiate blockers. Am J Ment Retard 1997; 102:182–199 Crossref, Medline, Google Scholar
76 : Beta-endorphin and ACTH are dissociated after self-injury in adults with developmental disabilities. Am J Ment Retard 2003; 108:414–424 Crossref, Medline, Google Scholar
77 : Self-injurious behavior and the efficacy of naltrexone treatment: a quantitative synthesis. Ment Retard Dev Disabil Res Rev 2004; 10:193–200 Crossref, Medline, Google Scholar
78 : Naltrexone in the treatment of dissociative symptoms in patients with borderline personality disorder: an open-label trial. J Clin Psychiatry 1999; 60:598–603 Crossref, Medline, Google Scholar
79 : Possible involvement of opioidergic and serotonergic mechanisms in antinociceptive effect of paroxetine in acute pain. J Pharmacol Sci 2004; 94:161–165 Crossref, Medline, Google Scholar
80 : Self-mutilation in personality disorders: psychological and biological correlates. Am J Psychiatry 1992; 149:221–226 Link, Google Scholar
81 : Evidence of endogenous opioid system dysregulation in borderline personality disorder. Biol Psychiatry 2008; 63:247S Google Scholar
82 : Single-nucleotide polymorphism in the human mu-opioid receptor gene alters beta-endorphin binding and activity: possible implications for opiate addiction. Proc Natl Acad Sci USA 1998; 95:9608–9613 Crossref, Medline, Google Scholar
83 : The single nucleotide polymorphism A118G alters functional properties of the human mu-opioid receptor. J Neurochem 2007; 103:77–87 Medline, Google Scholar
84 : Allelic expression imbalance of human mu-opioid receptor (OPRM1) caused by variant A118G. J Biol Chem 2005; 280:32618–32624 Crossref, Medline, Google Scholar
85 : Variation at the mu-opioid receptor gene (OPRM1) influences attachment behavior in infant primates. Proc Natl Acad Sci USA 2008; 105:5277–5281 Crossref, Medline, Google Scholar
86 : Opioid receptor and oxytocin genotypes in BPD, in 2009 Annual Meeting Syllabus and Proceedings Summary. Washington, DC, American Psychiatric Association, 2009 Google Scholar
87 : Opioid agonist efficacy predicts the magnitude of tolerance and the regulation of mu-opioid receptors and dynamin-2. Eur J Pharmacol 2007; 563:92–101 Crossref, Medline, Google Scholar
88 : Chronic opioid antagonist treatment dose-dependently regulates mu-opioid receptors and trafficking proteins in vivo. Pharmacol Biochem Behav 2003; 75:909–913 Crossref, Medline, Google Scholar
89 : Distribution of oxytocin and vasopressin in the rat supraoptic and paraventricular nucleus. J Endocrinol 1975; 67:461–462 Crossref, Medline, Google Scholar
90 : Multiple chemical messengers in hypothalamic magnocellular neurons. Prog Brain Res 1986; 68:161–168 Crossref, Medline, Google Scholar
91 : Variations in maternal behaviour are associated with differences in oxytocin receptor levels in the rat. J Neuroendocrinol 2000; 12:1145–1148 Crossref, Medline, Google Scholar
92 : Oxytocin and cortisol in romantically unattached young adults: associations with bonding and psychological distress. Psychophysiology 2008; 45:349–352 Crossref, Medline, Google Scholar
93 : Effects of partner support on resting oxytocin, cortisol, norepinephrine, and blood pressure before and after warm partner contact. Psychosom Med 2005; 67:531–538 Crossref, Medline, Google Scholar
94 : Oxytocin improves "mind-reading" in humans. Biol Psychiatry 2007; 61:731–733 Crossref, Medline, Google Scholar
95 : The integrative neurobiology of affiliation. Ann NY Acad Sci 1997; 807:xiii–xviii Crossref, Medline, Google Scholar
96 : Neuroendocrine mechanisms of stress and social interaction: implications for mental disorders. Curr Opin Psychiatry 2007; 20:158–162 Crossref, Medline, Google Scholar
97 : High doses of oxytocin cause sedation and low doses cause an anxiolytic-like effect in male rats. Pharmacol Biochem Behav 1994; 49:101–106 Crossref, Google Scholar
98 : Social support and oxytocin interact to suppress cortisol and subjective responses to psychosocial stress. Biol Psychiatry 2003; 54:1389–1398 Crossref, Medline, Google Scholar
99 : The role of CRF in behavioral aspects of stress. Ann NY Acad Sci 2001; 771:92–104 Crossref, Google Scholar
100 : Suckling-induced attenuation of plasma cortisol concentrations in postpartum lactating women. Endocr Res 1994; 20:79–87 Crossref, Medline, Google Scholar
101 : Neuropeptides and social behavior: animal models relevant to autism. Mol Psychiatry 2002; 7(suppl 2):S38–S39 Crossref, Medline, Google Scholar
102 : The neurobiology of love. FEBS Lett 2007; 581:2575–2579 Crossref, Medline, Google Scholar
103 : Neuroendocrine perspectives on social attachment and love. Psychoneuroendocrinology 1998; 23:779–818 Crossref, Medline, Google Scholar
104 : Evidence for a neuroendocrinological foundation of human affiliation: plasma oxytocin levels across pregnancy and the postpartum period predict mother-infant bonding. Psychol Sci 2007; 18:965–970 Crossref, Medline, Google Scholar
105 : Early experience in humans is associated with changes in neuropeptides critical for regulating social behavior. Proc Natl Acad Sci USA 2005; 102:17237–17240 Crossref, Medline, Google Scholar
106 : Sensitivity to intranasal oxytocin in adult men with early parental separation. Biol Psychiatry 2007; 61:1109–1111 Crossref, Medline, Google Scholar
107 : Lower CSF oxytocin concentrations in women with a history of childhood abuse. Mol Psychiatry 2009; 14:954–958 Crossref, Medline, Google Scholar
108 : Report of altered urinary oxytocin and AVP excretion in neglected orphans should be reconsidered. J Autism Dev Disord 2006; 36:829–830 Crossref, Medline, Google Scholar
109 : Oxytocin and social perception: oxytocin increases perceived facial trustworthiness and attractiveness. Horm Behav (in press) Google Scholar
110 : The neuroscience of affiliation: forging links between basic and clinical research on neuropeptides and social behavior. Horm Behav 2006; 50:518–528 Crossref, Medline, Google Scholar
111 : Oxytocin attenuates affective evaluations in conditioned faces and amygdala activity. J Neurosci 2008; 28:6607–6615 Crossref, Medline, Google Scholar
112 : Oxytocin enhances the encoding of positive social memories in humans. Biol Psychiatry 2008; 64:256–258 Crossref, Medline, Google Scholar
113 : Impact of oxytocin on stress-related dysphoria in borderline personality disorder (abstract). Biol Psychiatry 2009; 65(suppl 8):244S Google Scholar
114 : Effects of oxytocin in adults with borderline personality disorder and healthy adults (abstract). Biol Psychiatry 2009; 65(suppl 8):244S Google Scholar
115 : The rupture and repair of cooperation in borderline personality disorder. Science 2008; 312:806–810 Crossref, Google Scholar
116 : Vasopressin: behavioral roles of an "original" neuropeptide. Prog Neurobiol 2007; 84:1–24 Crossref, Medline, Google Scholar
117 : Oxytocin, vasopressin, and the neurogenetics of sociality. Science 2008; 322:900–904 Crossref, Medline, Google Scholar
118 : Vasopressin, oxytocin, and social behaviour. Curr Opin Neurobiol 2004; 14:777–783 Crossref, Medline, Google Scholar
119 : Paternal behavior influences development of aggression and vasopressin expression in male California mouse offspring. Horm Behav 2006; 50:699–707 Crossref, Medline, Google Scholar
120 : Enhanced partner preference in a promiscuous species by manipulating the expression of a single gene. Nature 2004; 429:754–757 Crossref, Medline, Google Scholar
121 : Microsatellite instability generates diversity in brain. Science 2005; 308:1630–1634 Crossref, Medline, Google Scholar
122 : Repeated agonistic encounters in hamsters modulate AVP V1a receptor binding. Horm Behav 2005; 48:545–551 Crossref, Medline, Google Scholar
123 : Vasopressin/serotonin interactions in the anterior hypothalamus control aggressive behavior in golden hamsters. J Neurosci 1997; 17:4331–4340 Crossref, Medline, Google Scholar
124 : A role for central vasopressin in pair bonding in monogamous prairie voles. Nature 1993; 365:545–548 Crossref, Medline, Google Scholar
125 : Central vasopressin administration regulates the onset of facultative paternal behavior in Microtus pennsylvanicus (meadow voles). Horm Behav 2001; 39:285–294 Crossref, Medline, Google Scholar
126 : Profound impairment in social recognition and reduction in anxiety-like behavior in vasopressin V1a receptor knockout mice. Neuropsychopharmacology 2004; 29:483–493 Crossref, Medline, Google Scholar
127 : Cerebrospinal fluid vasopressin levels: correlates with aggression and serotonin function in personality-disordered subjects. Arch Gen Psychiatry 1998; 55:708–714 Crossref, Medline, Google Scholar
128 : CSF biochemistries, glucose metabolism, and diurnal activity rhythms in alcoholic, violent offenders, fire setters, and healthy volunteers. Arch Gen Psychiatry 1994; 51:20–27 Crossref, Medline, Google Scholar
129 : Aggressive behavior linked to corticotrophin-reactive autoantibodies. Biol Psychiatry 2006; 60:799–802 Crossref, Medline, Google Scholar
130 : The effects of vasopressin on human facial responses related to social communication. Psychoneuroendocrinology 2004; 29:35–48 Crossref, Medline, Google Scholar
131 : Evidence for a dysfunctional prefrontal circuit in patients with an impulsive aggressive disorder. Proc Natl Acad Sci USA 2002; 99:8448–8453 Crossref, Medline, Google Scholar
132 : Arginine vasopressin and oxytocin modulate human social behavior. Ann NY Acad Sci 2009; 1167:87–102 Crossref, Medline, Google Scholar
133 : The oxytocin receptor (OXTR) contributes to prosocial fund allocations in the dictator game and the social value orientations task. PLoS ONE 2009; 4:e5535 Crossref, Medline, Google Scholar
134 : Association between arginine vasopressin 1a receptor (AVPR1a) promoter region polymorphisms and prepulse inhibition. Psychoneuroendocrinology 2009; 34:901–908 Crossref, Medline, Google Scholar
135 : Genetic variation in the vasopressin receptor 1a gene (AVPR1A) associates with pair-bonding behavior in humans. Proc Natl Acad Sci USA 2008; 105:14153–14156 Crossref, Medline, Google Scholar
136 : AVPR1A and OXTR polymorphisms are associated with sexual and reproductive behavioral phenotypes in humans. Hum Mutat 2007; 28:1150 Crossref, Medline, Google Scholar
137 : Mechanisms underlying epigenetic effects of early social experience: the role of neuropeptides and steroids. Neurosci Biobehav Rev 2005; 29:1089–1105 Crossref, Medline, Google Scholar
138 : Neonatal manipulations of oxytocin alter expression of oxytocin and vasopressin immunoreactive cells in the paraventricular nucleus of the hypothalamus in a gender-specific manner. Neuroscience 2004; 125:947–955 Crossref, Medline, Google Scholar
139 : Effects of early life stress on adult male aggression and hypothalamic vasopressin and serotonin. Eur J Neurosci 2006; 24:1711–1720 Crossref, Medline, Google Scholar
140 : Central vasopressin and oxytocin release: regulation of complex social behaviours. Prog Brain Res 2008; 170:261–276 Crossref, Medline, Google Scholar
141 : Carbetocin versus syntometrine in prevention of post-partum hemorrhage following vaginal delivery. J Obstet Gynaecol Res 2009; 35:48–54 Crossref, Medline, Google Scholar
142 : Clinical review: vasopression and terlipressin in septic shock patients. Crit Care 2005; 9:212–222 Crossref, Medline, Google Scholar
143 : Understanding and testing risk mechanisms for mental disorders. J Child Psychol Psychiatry 2008; 50:44–52 Crossref, Google Scholar
144 : Two-year randomized controlled trial and follow-up of dialectical behavior therapy vs therapy by experts for suicidal behaviors and borderline personality disorder. Arch Gen Psychiatry 2006; 63:757–766 Crossref, Medline, Google Scholar
145 : Evaluating three treatments for borderline personality disorder: a multiwave study. Am J Psychiatry 2007; 164:922–928 Link, Google Scholar
146 : 8-year follow-up of patients treated for borderline personality disorder: mentalization-based treatment versus treatment as usual. Am J Psychiatry 2008; 165:631–638 Link, Google Scholar
147 : Schema therapy for borderline personality disorder. J Clin Psychol 2006; 62:445–458 Crossref, Medline, Google Scholar
148 : Outpatient psychotherapy for borderline personality disorder: randomized trial of schema-focused therapy vs transference-focused psychotherapy. Arch Gen Psychiatry 2006; 63:649–658 Crossref, Medline, Google Scholar
149 : Vasopressin mediates enhanced offspring protection in multiparous rats. Neuropharmacology (Epub ahead of print, July 4, 2009) Google Scholar
150 : Neuropeptides and social behavior: effects of oxytocin and vasopressin in humans. Prog Brain Res 2008; 170:337–350 Crossref, Medline, Google Scholar
151 : Genetic variants in AVPR1A linked to autism predict amygdala activation and personality traits in healthy humans. Mol Psychiatry 2008; 133:1–8 Google Scholar
152 : Hypothalamic-pituitary-adrenocortical system dysregulation and new treatment strategies in depression. Expert Rev Neurother 2009; 9:1005–1019 Crossref, Medline, Google Scholar

