
Altered Brain Activation During Action Imitation and Observation in Schizophrenia: A Translational Approach to Investigating Social Dysfunction in Schizophrenia
Abstract
Objective
Social impairments are a key feature of schizophrenia, but their underlying mechanisms are poorly understood. Imitation, a process through which we understand the minds of others, involves the so-called mirror neuron system, a network comprising the inferior parietal lobe, inferior frontal gyrus, and posterior superior temporal sulcus. The authors examined mirror neuron system function in schizophrenia.
Method
Sixteen medicated schizophrenia patients and 16 healthy comparison subjects performed an action imitation/observation task during functional MRI. Participants saw a video of a moving hand or spatial cue and were instructed to either execute finger movements associated with the stimulus or simply observe. Activation in the mirror neuron system was measured during imitative versus nonimitative actions and observation of a moving hand versus a moving spatial cue. These contrasts were compared across groups.
Results
Activation in the mirror neuron system was less specific for imitation in schizophrenia. Relative to healthy subjects, patients had reduced activity in the posterior superior temporal sulcus during imitation and greater activity in the posterior superior temporal sulcus and inferior parietal lobe during nonimitative action. Patients also showed reduced activity in these regions during action observation. Mirror neuron system activation was related to symptom severity and social functioning in patients and to schizotypal syndrome in comparison subjects.
Conclusions
Given the role of the inferior parietal lobe and posterior superior temporal sulcus in imitation and social cognition, impaired imitative ability in schizophrenia may stem from faulty perception of biological motion and transformations from perception to action. These findings extend our understanding of social dysfunction in schizophrenia.
Social impairments are a central feature of schizophrenia (1), ranging from social withdrawal to misperceiving the intentions of others, with detrimental consequences for interpersonal relationships, quality of life, and functional outcomes. Moreover, social impairments are highly and uniquely predictive of conversion to psychosis in at-risk youths (2). Despite an increasing focus on understanding and treating these impairments, unraveling the faulty mental operations underlying social interactions in schizophrenia has proven difficult, arguably because currently available measures and tasks of social cognition are inadequate for isolating specific mechanisms.
Schizophrenia patients typically perform poorly on social cognitive tasks involving theory of mind (3), that is, tasks that require attributing mental states to oneself and others. However, theory-of-mind tasks often utilize complex stimuli that place a high demand on cognitive and perceptual functions that are compromised in schizophrenia. Thus, it is unclear whether the observed deficit reflects a specific social impairment or more generalized problems. An alternative approach to understanding social functioning in schizophrenia would be to examine basic building blocks that support higher-level social abilities.
Action imitation, the process by which one observes and replicates the actions of others, is one such building block of social cognition. Imitation ability is present from infancy (4) and is vital for nonverbal learning. Since social learning is almost entirely implicit and nonverbal in nature, imitation impairments can be expected to disproportionately affect social behavior. Imitation in the form of social mirroring facilitates communicative exchanges (5), and one hypothesized route toward understanding the minds of others is via covert imitative processes (6). That is, I understand your intentions and feelings by simulating my own experience to the same circumstances. Accordingly, imitation is considered the root of the ability to interpret the minds of others.
A further advantage to using imitation as a tool for studying social impairments is that neural correlates of action imitation are known at the cellular level. Neurophysiological studies of nonhuman primates have uncovered neurons, termed mirror neurons, in the premotor and parietal cortices that increase their firing rate when the monkey executes or observes an action. This mirror neuron system is hypothesized to form the neural basis of mirror matching and action imitation (Figure 1) (7). Within this framework, parietal neurons code for motoric components of the action and inferior frontal regions code for the action goal. Functional MRI (fMRI) studies have revealed an analogous network of regions involved in action imitation in humans (8). The posterior superior temporal sulcus, although it does not contain neurons with known mirror properties, is specialized for perceiving biological motion (9), which refers to the unique visual phenomenon of movements produced by living things; the posterior superior temporal sulcus provides this higher-order visual input to the mirror neuron system. Both temporoparietal and frontal regions contribute to the “core circuit” for imitation (7), and the mirror neuron system allows us to covertly simulate the sensation and movement of others.
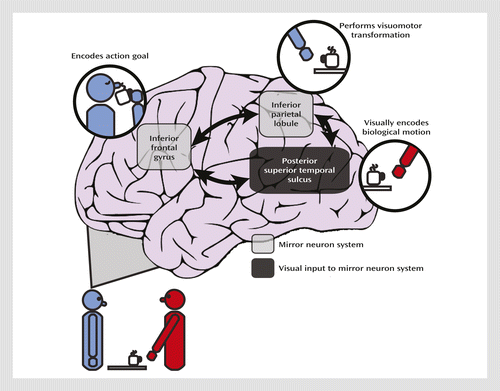
This translational approach, integrating paradigms adapted from primate neurophysiology and cognitive neuroscience, has been extremely valuable in understanding the etiology of cognitive impairments in schizophrenia (10). A similar strategy could be adopted for understanding social impairments in a wide range of psychiatric disorders. Indeed, imitation ability has been explored extensively in autism (11, 12) and is now a core part of behavioral therapies (13).
To date, studies of imitation in schizophrenia have been sparse, but impaired imitation has been reported in facial emotional expressions (14), as well as nonemotional facial and manual movements, which was found to be correlated with symptom severity (15, 16). Recent evidence also suggests deficient automatic mimicry in schizophrenia (17). Although no study has explicitly examined brain activity during action imitation in schizophrenia, abnormal posterior superior temporal activity during biological motion perception in schizophrenia has been reported (18).
Our goal in this fMRI study was to examine activation in the mirror neuron system of individuals with schizophrenia and healthy comparison subjects. Given findings of impaired imitation ability in schizophrenia, we hypothesized that we would find abnormal activity in the inferior frontal gyrus, the inferior parietal lobule, and the region around the posterior superior temporal sulcus and that it would be related to clinical symptoms and social functioning.
Method
Participants
The participants’ demographic and clinical characteristics are summarized in Table 1. Sixteen medicated outpatients with schizophrenia were recruited from a psychiatric facility in Nashville, Tenn. Diagnoses were made with the Structured Clinical Interview for DSM-IV. Symptoms were assessed with the Brief Psychiatric Rating Scale (BPRS) (19), the Scale for the Assessment of Positive Symptoms (20), and the Scale for the Assessment of Negative Symptoms (SANS) (21). Sixteen healthy participants with no history of DSM-IV axis I disorders were recruited from the same community by advertisements. Exclusion criteria for both groups were substance use or alcohol abuse within the past 6 months, brain injury, and neurological disease.
| Schizophrenia Patients (N=16) | Healthy Comparison Subjects (N=16) | |||||
|---|---|---|---|---|---|---|
| Characteristic | Mean | SD | Mean | SD | t | p |
| Age (years) | 40.2 | 9.1 | 37.4 | 7.0 | 1.1 | 0.29 |
| Handedness scoreb | 58.4 | 57.0 | 67.8 | 53.5 | 0.5 | 0.63 |
| Education (years) | 13.7 | 1.9 | 16.3 | 2.6 | 3.3 | 0.003 |
| Estimated IQc | 104.1 | 11.5 | 107.9 | 7.2 | 1.1 | 0.27 |
| Duration of illness (years) | 19.4 | 9.9 | ||||
| Scale for the Assessment of Positive Symptoms score | 14.1 | 9.8 | ||||
| Scale for the Assessment of Negative Symptoms score | 23.2 | 11.6 | ||||
| Brief Psychiatric Rating Scale score | 14.9 | 8.6 | ||||
| Schizotypal Personality Questionnaire | ||||||
| Total score | 10.8 | 6.6 | ||||
| Perceptual/cognitive subscore | 3.4 | 3.0 | ||||
| Interpersonal subscore | 5.9 | 4.5 | ||||
| Disorganized subscore | 2.6 | 2.3 | ||||
| Social Functioning Scale score | 128.9 | 21.2 | 158.3 | 12.1 | 4.8 | <0.001 |
Intelligence was estimated with the North American Adult Reading Test (22). Handedness was assessed using a modified Edinburgh Handedness Inventory (23). All participants had normal or corrected-to-normal vision. Groups were matched on age, sex, IQ, and handedness. Social functioning was assessed with the Social Functioning Scale (24). The Schizotypal Personality Questionnaire (25) was used to assess schizotypy in the healthy subjects. Participants gave written informed consent, as approved by the Vanderbilt Institutional Review Board, and they received compensation for their participation.
Experimental Paradigm
The fMRI task was adapted from previous studies of the mirror neuron system in healthy individuals (8). We modified it to require participants to press buttons on a button box for measurement of performance. A block design was used, with each 20-second block comprising three trials. Participants first saw an instructional cue to “execute” or “observe,” followed by a 2.5-second fixation period. Then, for each 3-second trial, participants saw one of three stimulus types (Figure 2). In the “animated” condition, a video of a hand pressing three buttons in sequence on a button box was presented. Under “execute” instructions, participants were asked to press the corresponding button simultaneously with the hand in the video. In the “symbolic” condition, participants were shown a still image of a hand on the same button box, with the cued fingers indicated with an X. Under “execute” instructions, participants were asked to press the button corresponding with the observed finger. Finally, in the “spatial” condition, three white X’s (placeholders) were shown, corresponding to finger positions, that would turn black to cue the appropriate finger. Under “execute” instructions, participants were asked to press the button corresponding to the cued position. Under “observe” instructions, participants simply watched the stimuli. Each trial contained three movements, and only the thumb, index, and middle finger were used. Each trial was followed by a 2.5-second fixation. Instruction and stimulus condition for each block were pseudorandomized. Twenty-second fixation blocks (randomly interleaved) served as a baseline. Participants performed four runs of 14 blocks with each hand; each run lasted 280 seconds. Accuracy and speed of key presses were recorded.
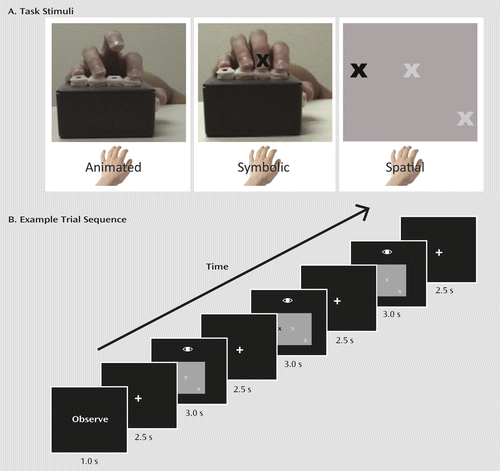
Visual stimuli, generated using MATLAB (MathWorks, Natick, Mass.) and the Psychophysics Toolbox (http://psychtoolbox.org), were back-projected onto a screen located at the observer’s feet and viewed through a periscope mirror attached to the head coil.
Data Acquisition
Images were acquired with a Philips Intera Achieva 3-T scanner. High-resolution T1 anatomical images were collected for each participant (170 slices; voxel size=1.0×1.0×1.0 mm3).Functional images were acquired parallel to the anterior commissure-posterior commissure line (gradient-echo T2*-weighted echo planar imaging sequence; 25 slices; TR=2000 ms, TE=35 ms, flip angle=79°, field of view=240×240 mm2, voxel size=1.875×1.875×5 mm3).
Data Analysis
Behavioral data.
Repeated-measures analyses of variance (ANOVAs) were conducted on the following variables: accuracy for the 1) execute trials and 2) the observe trials; 3) percentage of trials in which three movements were not completed during execute trials; response time to 4) initiate first movement and 5) complete movement sequence during execute trials. For each ANOVA, diagnostic group was entered as a between-subject variable and stimulus type as a within-subject variable.
fMRI data.
Data were preprocessed and analyzed using BrainVoyager QX1.10 (Brain Innovations, Maastricht, the Netherlands). Anatomical volumes were transformed into Talairach space on which functional volumes were aligned. Standard preprocessing was performed, including temporal high-pass frequency filtering, linear de-trending, three-dimensional motion correction, slice scan time correction, and spatial smoothing with a Gaussian kernel (6 mm full width at half maximum).
Individual subject analyses were performed using a general linear model with the six experimental conditions entered as block regressors, which were convolved with a canonical hemodynamic response function, and data were averaged across runs. First-level data were analyzed in a second-level random-effects analysis, using one-sample t tests to test the three contrasts of interest, outlined below. Independent t tests were used to measure group differences in activation related to these contrasts. Within- and between-group statistical maps were corrected for multiple comparisons at p<0.05 by using Monte Carlo simulations to arrive at a cluster extent threshold.
Our main contrasts of interest were imitation (execute-animated) > nonimitative action (execute-symbolic + execute-spatial), which isolated activity that was related to imitative action, and animated > nonanimated action observation, which isolated activity related to human movement observation (i.e., biological motion perception). Because there were no significant activation differences in any of our regions of interest between the execute-symbolic and execute-spatial conditions, nonimitative action activity was collapsed across these two conditions. Likewise, nonanimated action observation was collapsed across observe-symbolic and observe-spatial conditions.
To examine the relationship between brain activation and clinical status and social functioning in patients, correlational analyses were performed by extracting percent signal change from significant clusters of activation that were part of our three regions of interest and showed significant group differences in any of our contrasts of interest. Spearman’s rank correlation coefficients were calculated between the average percent signal change from each of these functionally defined regions and total scores on the Social Functioning Scale and the symptom assessment scales. For the healthy comparison subjects, percent signal change was extracted from the clusters of activation within the inferior frontal gyrus, inferior parietal lobe, and posterior superior temporal sulcus that showed greater activation for imitative than nonimitative action and correlated with Schizotypal Personality Questionnaire total and subscale scores.
To examine group differences in the amount of head motion, translational and rotational movement was averaged across runs and compared across groups.
Results
Detailed behavioral results are provided in the data supplement that accompanies the online edition of this article (see Behavioral Data and Table S1). Schizophrenia patients were less accurate than healthy comparison subjects in the execute condition, but mean accuracy for both groups was high (patients, 92%; healthy subjects, 97%), and there was no group difference in frequency of failure to complete a three-movement sequence. There was also no difference in accuracy in the observe condition. Thus, putative group differences in activation during execute trials cannot be explained by fewer movements in patients, nor can group differences in activation during observe trials be due to patients making errant key presses more frequently. Finally, collapsed across stimulus types, there was no group difference in latency to initiate the first movement or time to complete the movement sequence.
fMRI Data
Mean translational and rotational movement did not differ between groups. Table 2 lists coordinates of peak activation within our regions of interest for within- and between-group analyses (all between-group activation differences are listed in Table S2 in the online data supplement).
| Peak Talairach Coordinates | ||||||
|---|---|---|---|---|---|---|
| Contrast and Region | Cluster Size | x | y | z | Maximum t | Minimum p |
| Within groups | ||||||
| Imitation > nonimitative action execution | ||||||
| Healthy subjects | ||||||
| Right inferior parietal lobule/posterior superior temporal sulcus | 201 | 45 | –40 | 16 | 4.4 | 0.0001 |
| Right inferior frontal gyrus | 30 | 58 | 32 | 7 | 3.7 | 0.0009 |
| Schizophrenia patients | ||||||
| Right middle temporal gyrus/posterior superior temporal sulcus | 266 | 42 | –61 | 1 | 7.7 | <0.000001 |
| Left middle temporal gyrus/posterior superior temporal sulcus | 152 | –42 | –61 | 1 | 4.8 | 0.00004 |
| Animated > nonanimated action observation | ||||||
| Healthy subjects | ||||||
| Right postcentral gyrus/inferior parietal lobule | 56 | 48 | 35 | 16 | 3.9 | 0.0005 |
| Left inferior parietal lobule | 299 | –51 | –31 | 22 | 5.8 | 0.000002 |
| Schizophrenia patients | ||||||
| Right middle temporal gyrus/posterior superior temporal sulcus | 149 | 39 | –61 | 1 | 6.6 | <0.000001 |
| Left middle temporal gyrus/posterior superior temporal sulcus | 95 | –45 | –61 | –2 | 4.8 | 0.00004 |
| Between groups | ||||||
| Imitation > nonimitative action execution | ||||||
| Healthy subjects > schizophrenia patients | ||||||
| Right inferior parietal lobule/posterior superior temporal sulcus | 94 | 45 | –40 | 22 | 4.5 | 0.00008 |
| Imitation > baseline | ||||||
| Healthy subjects > schizophrenia patients | ||||||
| Right posterior superior temporal sulcus | 63 | 45 | –43 | 19 | 4.2 | 0.0002 |
| Left inferior parietal lobule | 13 | –48 | –34 | 31 | 3.6 | 0.001 |
| Nonimitative action execution > baseline | ||||||
| Schizophrenia patients > healthy subjects | ||||||
| Right inferior parietal lobule | 512 | 51 | –34 | 22 | 5.0 | 0.00003 |
| Left posterior superior temporal sulcus | 135 | –39 | –28 | 4 | 3.8 | 0.0007 |
| Animated > nonanimated action observation | ||||||
| Healthy subjects > schizophrenia patients | ||||||
| Right inferior parietal lobule | 9 | 51 | –25 | 22 | 3.5 | 0.002 |
| Animated action observation > baseline | ||||||
| Healthy subjects > schizophrenia patients | ||||||
| Right inferior parietal lobule | 184 | 51 | –28 | 22 | 4.5 | 0.00009 |
| Right inferior parietal lobule/posterior superior temporal sulcus | 61 | –43 | 22 | 4.0 | 0.0004 | |
| Left posterior superior temporal sulcus | 33 | –42 | –31 | 13 | 3.5 | 0.001 |
Imitation versus nonimitative action.
Consistent with previous studies, healthy subjects activated the mirror neuron system and showed greater activation in the posterior superior temporal sulcus, inferior parietal lobe, and inferior frontal gyrus for imitative compared with nonimitative action. Activation reached significance only in the right hemisphere. Patients also showed bilateral activation in the inferior parietal lobe and posterior superior temporal sulcus. Direct group comparison revealed a more differentiated response to imitative compared with nonimitative action in healthy subjects in a region that was maximally different in the right inferior parietal lobe and extended into the posterior superior temporal sulcus (Figure 3). The basis of this difference was greater activation in healthy subjects than in patients in the posterior superior temporal sulcus for imitative actions, but greater activation in patients than in healthy subjects in the right inferior parietal lobe and posterior superior temporal sulcus for nonimitative action. Healthy subjects also showed greater activation to imitative action than patients in a second, more anterior, region of the right posterior superior temporal sulcus and in the left inferior parietal lobe, and patients showed greater activation to nonimitative action than healthy subjects in the left posterior superior temporal sulcus.
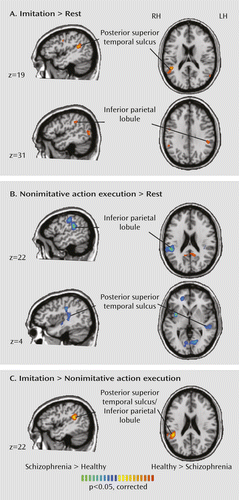
a BOLD=blood-oxygen-level-dependent.
Animated versus nonanimated action observation.
Compared with patients, healthy subjects showed a greater differentiated response to animated relative to nonanimated action observation in the right inferior parietal lobe (Figure 4). The basis of this difference was greater activation in the right inferior parietal lobe extending into the posterior superior temporal sulcus in healthy subjects for animated action observation. Additionally, healthy subjects had greater activation for observing animated actions in the left posterior superior temporal sulcus, indicating a robust response to biological motion. There were no significant group differences in activation patterns in any of our regions of interest during observation of nonanimated actions.
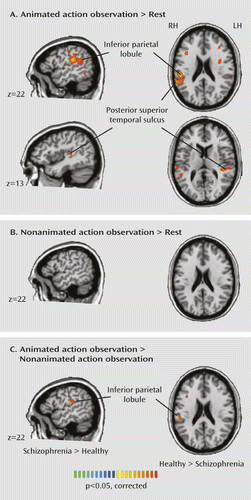
a BOLD=blood-oxygen-level-dependent.
Correlational analyses.
In patients, greater activity in the right posterior superior temporal sulcus during action imitation was related to lower BPRS score (rs=−0.50, p=0.051). Additionally, greater activity in the right inferior parietal lobe during nonimitative action execution was related to lower SANS score (rs=−0.48, p=0.054). Somewhat counterintuitively, greater activation in the right inferior parietal lobe for animated action observation than nonanimated action observation was related to poorer Social Functioning Scale score (rs=−0.57, p=0.02). Higher antipsychotic dosage was related to greater activity in the right inferior parietal lobe for animated versus nonanimated action observation (rs=0.54, p=0.03).
Finally, in healthy subjects, psychometric schizotypy was associated with imitation-related brain activation, but the association fell short of statistical significance; greater activation in the right inferior frontal gyrus for imitative compared with nonimitative action was associated with higher score on the Schizotypal Personality Questionnaire (r=0.43, p=0.09), driven by the interpersonal subscale score (r=0.41, p=0.06). Scatterplots are presented in Figure S1 in the online data supplement.
Discussion
Our results indicate abnormal neural activity during action imitation and observation in patients with schizophrenia. Instead of a simple reduction in mirror neuron system activation in schizophrenia, we observed an interesting crossover effect. Patients showed a less differentiated response than healthy subjects in the inferior parietal lobe and posterior superior temporal sulcus during action imitation. Although patients had less activation than healthy subjects in the posterior superior temporal sulcus during imitation, they had greater activation in the posterior superior temporal sulcus and inferior parietal lobe during nonimitative action, suggesting that the mirror neuron system is less fine-tuned in schizophrenia. Patients also showed reduced activation in the inferior parietal and posterior superior temporal regions during observation of a moving hand.
In interpreting the significance of these findings, we turn to accounts of posterior superior temporal sulcus and inferior parietal lobe function in the healthy brain. The posterior superior temporal sulcus plays a crucial role in biological motion perception (9, 26) and provides higher-order visual input necessary for imitation to the mirror neuron system. Accordingly, abnormal activation in this region during imitation and observation in schizophrenia suggests that impaired imitation (14–16) may have its basis largely in faulty perceptual input to brain regions more directly implicated in imitation. Our results bear a striking similarity to earlier findings (18) showing abnormal posterior superior temporal sulcus activity during biological motion perception in schizophrenia. In that study, patients tended to perceive randomly moving dots as having biological attributes. Failure to discriminate biological from nonbiological motion in schizophrenia was reflected in the posterior superior temporal sulcus, with no differential activity in this region between biological and nonbiological motion perception.
We also observed abnormal activation in patients in the nearby inferior parietal lobe, which is densely interconnected to the posterior superior temporal sulcus (27). Based on the involvement of the inferior parietal lobe in action imitation and observation (7), apraxia resulting from inferior parietal lobe lesions (28), and the multimodal nature of representations stored in the inferior parietal lobe (29), a unique role for this region has been postulated. The inferior parietal lobe is argued to be involved in matching the perceptual information about an observed action with a motoric representation that action; this function gives rise to the ability to understand actions, and ultimately, intentions. This claim is bolstered by fMRI findings of involvement of the temporoparietal junction, comprising the inferior parietal lobe and superior temporal regions, during more complex theory-of-mind tasks (30). We observed greater activity in the right inferior parietal lobe during nonimitative actions and reduced activity in the inferior parietal lobe during imitative actions and action observation in patients, which is suggestive of reduced fine-tuning of this region for socially relevant stimuli in schizophrenia.
Mounting evidence implicates inferior parietal dysfunction in schizophrenia. In a review, Torrey (31) highlighted the late development of this region, both evolutionarily and over the lifespan, which is consistent with the late onset of the disease and the notion that schizophrenia is a uniquely human disorder (32). Moreover, the inferior parietal lobe is crucial for self-related processes (e.g., perception of agency) central to illness phenomenology. Inferior parietal lobe hyperactivity has been observed in multiple cognitive and sensorimotor tasks in schizophrenia (see reference 31 for a review). Furthermore, the most robust functional connectivity abnormalities in schizophrenia across the brain occur between parietal nodes, in which increased connectivity has been reported (33). Our findings are consistent with abnormal parietal lobe function in schizophrenia.
Increasing evidence suggests impaired imitation in schizophrenia, but what implications do these findings have for understanding the clinical picture, and how do our findings of abnormal neural activity during action imitation elucidate the underlying pathology? One route to understanding the minds of others is via internalized mimicking. Additionally, automatic mimicry enhances the subjective quality of social interactions (5). One might postulate that disturbances in imitation could cascade into a failure to understand the minds of others (16) and that poor social interactions could fuel social isolation, which in turn can worsen symptoms (34).
Given the purported role of the inferior parietal lobe and posterior superior temporal sulcus as key nodes in an imitation network, findings of abnormal activity in these regions in schizophrenia suggest that imitation impairments may have their basis in faulty biological motion perception and transformation of visual information into motor representations. Hyperactivity in these regions during nonimitative action execution and hypoactivation of these regions during imitation generates interesting hypotheses about the nature of social impairments in schizophrenia. The combination of hyper- and hypoactivation in these regions may correspond to the clinical presentation of social dysfunction in schizophrenia, which is characterized by both enhanced sensitivity to social information (e.g., suspiciousness about others, perceiving animacy and intentions in inanimate objects) and reduced sensitivity to the states of others (e.g., social withdrawal, avolition). We observed correlations that bordered on significance between symptom severity and neural activity in the posterior superior temporal sulcus and inferior parietal lobe during both imitative and nonimitative actions. It should be noted, however, that given the exploratory nature of these correlations, they were not corrected for multiple comparisons, and larger and more clinically heterogeneous samples are required to fully explore this link.
One important question for future study is the developmental origin of imitation impairments in schizophrenia. One possibility is that imitation ability fails to develop appropriately. Another is that mirror-matching abilities break down during illness onset. It is known that experience in the form of sensorimotor learning can mediate the automaticity of imitation (35). If the link between motor commands and sensory consequences becomes degraded in schizophrenia, perhaps via alterations in corollary discharge (36), one could envision a gradual breakdown in mirror-matching. Furthermore, dysfunctional simulation of the intentions of others would spur the need for alternative sources of information, perhaps through prior knowledge. Indeed, Chambon et al. (37) found that schizophrenia patients were more likely to use prior expectations over current information when deciphering the actions of others. While this notion is speculative, it forms a basis for further inquiry.
Although inferior frontal activation did not significantly differ between healthy subjects and schizophrenia patients for any of the contrasts of interest, we observed a significant correlation between inferior frontal activation and schizotypal traits in healthy participants. Score on the interpersonal subscale of the Schizotypal Personality Questionnaire was associated with increased inferior frontal activity during imitation. These results suggest that healthy individuals with elevated schizotypy may be expending greater effort to correctly imitate the actions of others. Platek et al. (38) also observed subtle imitative abnormalities in relation to elevated schizotypy.
There are several limitations to our study. First, the patients were medicated. Although the effects of antipsychotic medication on imitation and action observation are unknown, antipsychotic dosage was related to inferior parietal lobe activity in the imitation versus nonimitative action contrast. That is, brain activity in patients with higher medication dosages looked more like that of healthy subjects, suggesting a possible therapeutic effect of antipsychotic treatment. Second, there were subtle group differences in task performance; however, accuracy was high in both groups, response time differences were small, and there was no difference in the total number of movements made, suggesting that differences in brain activation cannot be explained by the amount of motor output. Finally, we did not independently measure attention during action observation. However, the brain activity during observe trials was markedly different from that during rest blocks, and groups were matched on IQ, which precludes the possibility of gross differences in cognitive functioning.
Conclusions
We observed abnormal activity in the mirror neuron system in schizophrenia. In patients, activation in the posterior superior temporal sulcus and inferior parietal lobe was less specific to action imitation compared with nonimitative action. Furthermore, activity in these two regions was reduced during action observation in schizophrenia. Given the specific role of these regions in social cognition, impaired imitative ability may be rooted in faulty perception of biological motion and erroneous transformation of visual information into motor representations. These findings have significant implications for understanding the nature of social dysfunction in schizophrenia.
1 : Social cognition in schizophrenia: an NIMH workshop on definitions, assessment, and research opportunities. Schizophr Bull 2008; 34:1211–1220Crossref, Medline, Google Scholar
2 : Prediction of psychosis in youth at high clinical risk: a multisite longitudinal study in North America. Arch Gen Psychiatry 2008; 65:28–37Crossref, Medline, Google Scholar
3 : “Theory of mind” in schizophrenia: a review of the literature. Schizophr Bull 2005; 31:21–42Crossref, Medline, Google Scholar
4 : Imitation of facial and manual gestures by human neonates. Science 1977; 198:75–78Crossref, Medline, Google Scholar
5 : The chameleon effect: the perception-behavior link and social interaction. J Pers Soc Psychol 1999; 76:893–910Crossref, Medline, Google Scholar
6 Carruthers PSmith PK (eds): Theories of Theory of Mind. Cambridge, UK, Cambridge University Press, 1996Crossref, Google Scholar
7 : Neural mechanisms of imitation. Curr Opin Neurobiol 2005; 15:632–637Crossref, Medline, Google Scholar
8 : Cortical mechanisms of human imitation. Science 1999; 286:2526–2528Crossref, Medline, Google Scholar
9 : Brain areas active during visual perception of biological motion. Neuron 2002; 35:1167–1175Crossref, Medline, Google Scholar
10 : Identifying cognitive mechanisms targeted for treatment development in schizophrenia: an overview of the first meeting of the Cognitive Neuroscience Treatment Research to Improve Cognition in Schizophrenia Initiative. Biol Psychiatry 2008; 64:4–10Crossref, Medline, Google Scholar
11 : Imitation and action in autism: a critical review. Psychol Bull 1994; 116:259–273Crossref, Medline, Google Scholar
12 : Neural mechanisms of imitation and “mirror neuron” functioning in autistic spectrum disorder. Neuropsychologia 2006; 44:610–621Crossref, Medline, Google Scholar
13 : Teaching reciprocal imitation skills to young children with autism using a naturalistic behavioral approach: effects on language, pretend play, and joint attention. J Autism Dev Disord 2006; 36:487–505Crossref, Medline, Google Scholar
14 : Imitation of facial expressions in schizophrenia. Psychiatry Res 2006; 145:87–94Crossref, Medline, Google Scholar
15 : Gesture imitation in schizophrenia. Schizophr Bull 2013; 39:94–101Crossref, Medline, Google Scholar
16 : Imitation, simulation, and schizophrenia. Schizophr Bull 2008; 34:698–707Crossref, Medline, Google Scholar
17 : Empathic deficits in schizophrenia: the potential role of rapid facial mimicry. J Int Neuropsychol Soc 2010; 16:621–629Crossref, Medline, Google Scholar
18 : Perception of biological motion in schizophrenia and healthy individuals: a behavioral and FMRI study. PLoS ONE 2011; 6:e19971Crossref, Medline, Google Scholar
19 : The Brief Psychiatric Rating Scale. Psychol Rep 1962; 10:799–812Crossref, Google Scholar
20 : Scale for the Assessment of Positive Symptoms (SAPS). Iowa City, University of Iowa, 1984Google Scholar
21 : Scale for the Assessment of Negative Symptoms (SANS). Iowa City, University of Iowa, 1983Google Scholar
22 : Predicting premorbid IQ: a revision of the national Adult Reading Test. Clin Neuropsychol 1989; 3:129136Crossref, Google Scholar
23 : The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 1971; 9:97–113Crossref, Medline, Google Scholar
24 : The Social Functioning Scale: the development and validation of a new scale of social adjustment for use in family intervention programmes with schizophrenic patients. Br J Psychiatry 1990; 157:853–859Crossref, Medline, Google Scholar
25 : The SPQ: a scale for the assessment of schizotypal personality based on DSM-III-R criteria. Schizophr Bull 1991; 17:555–564Crossref, Medline, Google Scholar
26 : Action observation circuits in the macaque monkey cortex. J Neurosci 2011; 31:3743–3756Crossref, Medline, Google Scholar
27 : Common cortical and subcortical targets of the dorsolateral prefrontal and posterior parietal cortices in the rhesus monkey: evidence for a distributed neural network subserving spatially guided behavior. J Neurosci 1988; 8:4049–4068Crossref, Medline, Google Scholar
28 : Deconstructing apraxia: understanding disorders of intentional movement after stroke. Curr Opin Neurol 2002; 15:71–77Crossref, Medline, Google Scholar
29 : Spatial coding of visual and somatic sensory information in body-centred coordinates. Eur J Neurosci 2001; 14:737–746Crossref, Medline, Google Scholar
30 : Neuronal correlates of theory of mind and empathy: a functional magnetic resonance imaging study in a nonverbal task. Neuroimage 2006; 29:90–98Crossref, Medline, Google Scholar
31 : Schizophrenia and the inferior parietal lobule. Schizophr Res 2007; 97:215–225Crossref, Medline, Google Scholar
32 : Is schizophrenia the price that Homo sapiens pays for language? Schizophr Res 1997; 28:127–141Crossref, Medline, Google Scholar
33 : Key functional circuitry altered in schizophrenia involves parietal regions associated with sense of self. Hum Brain Mapp 2014; 35:123–139Crossref, Medline, Google Scholar
34 : A social deafferentation hypothesis for induction of active schizophrenia. Schizophr Bull 2007; 33:1066–1070Crossref, Medline, Google Scholar
35 : Associative sequence learning: the role of experience in the development of imitation and the mirror system. Philos Trans R Soc Lond B Biol Sci 2009; 364:2369–2380Crossref, Medline, Google Scholar
36 : Efference copy and corollary discharge: implications for thinking and its disorders. Schizophr Bull 1978; 4:636–640Crossref, Medline, Google Scholar
37 : Mentalizing under influence: abnormal dependence on prior expectations in patients with schizophrenia. Brain 2011; 134:3728–3741Crossref, Medline, Google Scholar
38 : Contagious yawning: the role of self-awareness and mental state attribution. Brain Res Cogn Brain Res 2003; 17:223–227Crossref, Medline, Google Scholar

