
Neuroanatomic Alterations and Social and Communication Deficits in Monozygotic Twins Discordant for Autism Disorder
Abstract
Objective: Investigating neuroanatomic differences in monozygotic twins who are discordant for autism can help unravel the relative contributions of genetics and environment to this pervasive developmental disorder. The authors used magnetic resonance imaging (MRI) to investigate several brain regions of interest in monozygotic twins who varied in degree of phenotypic discordance for narrowly defined autism. Method: The subjects were 14 pairs of monozygotic twins between the ages of 5 and 14 years old and 14 singleton age- and gender-matched typically developing comparison subjects. The monozygotic twin group was a cohort of children with narrowly defined autistic deficits and their co-twins who presented with varying levels of autistic deficits. High-resolution MRIs were acquired and volumetric/area measurements obtained for the frontal lobe, amygdala, and hippocampus and subregions of the prefrontal cortex, corpus callosum, and cerebellar vermis. Results: No neurovolumetric/area differences were found between twin pairs. Relative to typically developing comparison subjects, dorsolateral prefrontal cortex volumes and anterior areas of the corpus callosum were significantly altered in autistic twins, and volumes of the posterior vermis were altered in both autistic twins and co-twins. Intraclass correlation analysis of brain volumes between children with autism and their co-twins indicated that the degree of within-pair neuroanatomic concordance varied with brain region. In the group of subjects with narrowly defined autism only, dorsolateral prefrontal cortex, amygdala, and posterior vermis volumes were significantly associated with the severity of autism based on scores from the Autism Diagnostic Observation Schedule—Generic. Conclusions: These findings support previous research demonstrating alterations in the prefrontal cortex, corpus callosum, and posterior vermis in children with autism and further suggest that alterations are associated with the severity of the autism phenotype. Continued research involving twins who are concordant and discordant for autism is essential to disentangle the genetic and environmental contributions to autism.
Autism is defined by the presence of markedly abnormal development in social interaction, communication, and repertoire of activity and interests (DSM-IV). The dynamic neurological and social deficits observed in children with autism contribute to varying levels of impairment. Although advancements in autism research have improved our understanding of the disorder, many questions regarding the pathophysiology of autism remain (1) .
Two decades of magnetic resonance imaging (MRI) studies have yielded somewhat consistent findings of specific brain regions that are altered in children with autism, including the frontal lobe (2 – 4) , amygdala (1 , 5 – 11) , cerebellar vermis (12 – 21) , and corpus callosum (14 , 22 – 27) .
Studies indicate that young children with autism initially exhibit brain enlargement, followed by a decrease in overall brain volume over time. Brain enlargement is most pronounced in the frontal lobe (28) , possibly as a result of the late development of this region (29) . However, the specific neuroanatomic subregions that contribute to growth dysregulation of the frontal lobe have not been firmly established. Enlargement has been observed in dorsolateral and medial frontal regions (2) but not in the orbitofrontal cortex (3) of children with autism. It has been suggested that possible alterations in neuronal development of the frontal cortex in autism ultimately may lead to abnormal local and long-distance neural connectivity (29) . Supporting this hypothesis, in part, are diffusion tensor and anatomic imaging studies of the corpus callosum, in which reductions in fractional anisotrophy and/or volumes have been observed in both anterior and posterior aspects of this structure in individuals with autism (23 – 26 , 30 , 31) .
Less consistent are MRI studies of the cerebellar vermis in autism, some of which have observed vermal hypoplasia (12) , particularly in lobules VI and VII, and others have observed vermal hyperplasia (13) . Variability in findings are most likely attributable to the heterogeneity of the disorder (13) .
Similarly, studies of alterations of the amygdala in autism have been inconsistent. Results vary between increased (7 , 9 , 32) , decreased (5 , 6) , and same-sized amygdala volumes (33) . One possible explanation for these seemingly contradictory findings is the variability in age and degree of severity across autistic subject populations. Further, a severity-by-age interaction has been proposed, suggesting that younger less affected autistic subjects would have normal to enlarged amygdala volumes, while older more affected individuals would have smaller amygdala volumes (10) .
Several lines of research suggest that genetic, epigenetic, and environmental interactions structure the brain (34) and therefore contribute to autistic deficits as well as accompanying neuroanatomic anomalies. Studying monozygotic twins with autism who may be discordant for the disorder can elucidate the respective influences of genetic and environmental factors on susceptibility to autism. In a previous study of neuroanatomic variation in a cohort of monozygotic twin pairs discordant for the narrow phenotype for autism (4) , we used a semiautomated method to measure CSF and gray and white tissue compartments of lobar regions. We found that relative to typically developing comparison subjects, both children with autism and their co-twins exhibited decreased volumes in frontal, temporal, and occipital white matter. In order to extend the findings of our previous study, which did not focus on sublobar regions, we conducted manual volumetric measurements of the total frontal and prefrontal cortices, corpus callosum, amygdala, hippocampus, and cerebellar vermis in this cohort. Based on previous research, we hypothesized that 1) volumes of the dorsolateral and dorsomedial cortical subregions and the amygdala would be enlarged; 2) areas/volumes of the corpus callosum and lobules VI and VII of the cerebellar vermis would be decreased in children with autism (although not necessarily their co-twins); and 3) the volumes of those regions that significantly differ between children with autism and comparison subjects would be associated with behavioral measures of severity of autism.
Previous neuroanatomic studies of typically developing monozygotic twins suggest a high degree of concordance in brain volumes. Giedd and colleagues (35) performed multivariate analyses on neuroanatomic brain volumes of pediatric monozygotic and same-sex dizygotic twin pairs with typical development. They observed within-structure cross-twin correlations that were substantially higher for monozygotic twins relative to dizygotic twins. A pediatric imaging study of monozygotic and dizygotic twins conducted by Wallace and colleagues (36) reported monozygotic correlations of gray and white matter volumes from different neuroanatomic regions of the brain that were nearly twice as large as dizygotic volumetric correlations of the same regions. In our previous study, we found that cerebral, but not cerebellar, gray and white matter volumes were highly correlated for both clinically concordant and discordant twin pairs. Considering this evidence, we would expect this same cohort to have highly concordant frontal lobe, corpus callosum, hippocampus, and amygdala volumes but not highly concordant cerebellar vermis volumes, suggesting a higher degree of genetic control over cerebral development relative to cerebellar development.
Method
Research Subjects
Participants were 42 children between the ages of 5 and 14 years old (14 pairs of monozygotic twins and 14 age- and gender-matched typically developing comparison subjects). The mean age of the twin pairs was 8.8 years (SD=2.6 years; range=5.6–13.8). The mean age of the comparison subjects was 8.4 years (SD=2.6 years; range=5.4–13.9). Twelve twin pairs and their comparison subjects were boys.
Families of children with autism were recruited through the Autism Society of America, the National Alliance for Autism Research, the Kennedy Krieger Center for Autism and Related Disorders, and clinical neurologists at the Kennedy Krieger Institute. Recruitment methods are described in detail elsewhere (4) . In order to qualify for a diagnosis of autism, a child had to meet criteria for autism on the Autism Diagnostic Interview—Revised (37) and score within one point of the criteria for autism on the Autism Diagnostic Observation Schedule—Generic (38) . Each monozygotic twin pair in the study consisted of at least one child with a diagnosis of autism based on these criteria.
DNA fingerprinting probes were used to confirm zygosity in all twin pairs. Eight independent loci were tested for each twin pair. The DNA profiles for all twin pairs were statistically identical at every locus, indicating that the probability of monozygosity for each twin pair was approximately 99.99%.
Typically developing singleton comparison subjects were recruited from the office of a local pediatrician. Developmental diagnoses, which included language delay and psychiatric diagnoses (e.g., attention deficit hyperactivity disorder), were ruled out by parental report and the Child Behavior Checklist (39) . None of the comparison subjects had known neurologic, developmental, learning, or psychiatric disorders.
MRI Pulse Sequence
MRI was performed on the same 1.5 Tesla GE Signa scanner (General Electric, Milwaukee). A three-dimensional high-resolution MRI scan was acquired for each subject using a T1-weighted spoiled gradient-echo sequence (time to repeat=35–45 msec, echo time=5–7 msec, number of excitations=1, flip angle=45°, matrix size=256×128, field of view=20–24 mm, yield=124 coronal slices, slice thickness=1.5 mm).
Measurement Protocols
Images were imported into the imaging software program BrainImage (40) to remove nonbrain tissue and to measure the amygdala, hippocampus, and corpus callosum. Subsequently, images were moved to the software program Measure for measurement of the frontal lobe, subregions of the prefrontal cortex (dorsolateral, dorsomedial, orbitolateral, and orbitomedial), and the cerebellar vermis. Measures of all brain regions were based on previously published protocols (41 – 45) . Raters were blind to diagnosis. Reliability coefficients were calculated for all regions of interest with the intraclass correlation coefficient, and raters achieved coefficients ranging from 0.90 to 0.99 for the regions measured.
Autism Diagnostic Interviews
The Autism Diagnostic Interview—Revised was administered by telephone during separate interviews for each twin by a staff member trained in the reliable administration of this instrument. The Autism Diagnostic Observation Schedule—Generic was administered by the same staff member at our center. Scores were used to diagnose and determine the severity of autism in each subject.
Demographic Variables
In order to further characterize our subjects, the Stanford-Binet Intelligence Scale—Fourth Edition (46) was administered to the autism and co-twin groups, and the WISC-III (47) was administered to the comparison group. The comparison group was administered a different IQ test because they were acquired simultaneously for a separate study that necessitated the use of the WISC-III. Socioeconomic status was measured using the Two-Factor Index of Social Standing (48) .
Statistical Analyses
Analyses of variance (ANOVAs) assessed the effect of study group on volumes of total frontal lobe. Analyses of covariance (ANCOVAs) assessed the effect of study group on volumes of the amygdala and hippocampus, with whole brain volume entered as a covariate. Multiple analyses of covariance (MANCOVAs) assessed the effect of study group on 1) total dorsolateral prefrontal cortex, dorsomedial prefrontal cortex, orbitolateral prefrontal cortex, and orbitomedial prefrontal cortex volumes; 2) the five subregions of the corpus callosum; and 3) the three subregions of the cerebellar vermis. Whole brain volume was entered as a covariate in these analyses. Effect size (ε 2 ) is provided for all analyses of main effects. Intraclass correlation coefficients were used to assess concordance in volumes of regions of interest between twin pairs. Hierarchical regression analyses were conducted in order to determine associations between brain volumes and total scores on both autism diagnostic scales. Since the twin pairs differed significantly in cognitive levels, full-scale IQ was entered into the model initially, followed by volumes of brain regions of interest, all of which were entered as a block.
Results
Sample Characteristics
There were no significant differences between our autism subjects, their discordant co-twins, and the unrelated matched comparison subjects in terms of age or socioeconomic status. All groups differed significantly in IQ scores, as shown in Table 1 . In terms of categorical levels of intellectual functioning, 62% of participants with autism had IQ scores in the range of mental retardation, 15% in the borderline range, and 23% in the average range. In contrast, 23% of IQ scores among co-twins fell in the range of mental retardation, 23% in the borderline range, and 54% in the average range. Between-twin comparisons of standard subscale scores on the Stanford-Binet Intelligence Scale ( Table 1 ) indicated that twin pairs differed significantly in verbal reasoning (p=0.04), abstract/visual reasoning (p=0.02), and quantitative reasoning (p=0.04) but not short-term memory (p=0.12).

Autism and co-twin groups differed significantly in total scores on both the Autism Diagnostic Interview (F=16.11, df=1, 25, p=0.001) and the Autism Diagnostic Observation Schedule (F=20.75, df=1, 25, p<0.001) ( Table 2 ). Thirty-six percent of the co-twins were diagnosed with autism and therefore were clinically concordant with their twin for autism. Twenty-eight percent of co-twins were diagnosed with pervasive developmental disorder, and 36% had no diagnosis.

Volumetric Comparisons
The volumetric means of all regions of interest are shown in Table 3 . ANOVAs indicated that although children with autism and their co-twins displayed smaller absolute frontal lobe volumes relative to comparison subjects the differences were not significant (F=1.43, df=2, 39, p=0.25), even after covarying for whole brain volume (p=0.28).

A MANCOVA assessing the effects of study group on all prefrontal subregions yielded a Wilks’ lambda distribution of 0.55 (p=0.009). Only dorsolateral prefrontal cortex volumes differed between study groups (F=4.52, df=2, 36, p=0.02; ε 2 =0.20). Whole brain volume was also a significant covariate (p=0.045). Follow-up ANOVAs indicated that the right (but not left) dorsolateral prefrontal cortical volumes for children with autism were significantly larger (F=4.61, df=2, 39, p=0.02; ε 2 =0.191) than those of comparison subjects (p=0.01) but not co-twins (p=0.25).
Corpus callosum areas differed significantly between study groups, yielding a Wilks’ lambda distribution of 0.58 (p=0.047). Only the anterior body and the genu accounted significantly for these differences. Whole brain volume was also a significant covariate (p=0.005). Univariate ANOVAs indicated that genu (F=3.19, df=2, 36, p=0.05; ε 2 =0.15) and anterior body (F=4.18, df=2, 36, p=0.02; ε 2 =0.19) areas were smaller for children with autism relative to comparison subjects but not to co-twins.
The omnibus model for cerebellar vermis volumes was not significant (Wilks’ lambda distribution=0.77, p=0.17). However, univariate analyses indicated that lobules VI and VII differed between study groups (F=3.54, df=2, 38, p=0.04; ε 2 =0.17). Whole brain volume was also a significant covariate. Planned comparisons indicated that relative to comparison subjects lobules VI and VII were significantly smaller in co-twins (p=0.02) and tended to be smaller in children with autism (p=0.06). Amygdala and hippocampus volumes did not differ significantly between study groups.
Neuroanatomic Concordance Between Twin Pairs
Neuroanatomic concordance between twin pairs was assessed with the intraclass correlation coefficient and yielded the following results: whole brain volume, 0.93; frontal lobe, 0.93; dorsolateral prefrontal cortex, 0.65; corpus callosum, 0.78; amygdala, 0.66; hippocampus, 0.91; and cerebellar vermis, 0.82.
Association Between Neuroanatomic Volumes and Severity of Autism
Neuroanatomic regions that significantly differentiated study groups were included in hierarchical regression analyses, including the right dorsolateral prefrontal volumes, anterior body area of the corpus callosum, and vermis lobules VI and VII volumes. Amygdala volumes were also included, since previous studies have reported an association with social and communication deficits in autism (1 , 49) . Table 4 summarizes these results.
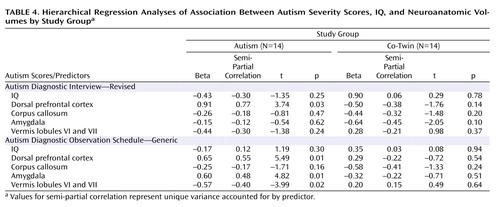
When measured with the Autism Diagnostic Interview, autism severity was not significantly associated with either IQ scores or the block of neuroanatomic volumes for twins with autism or their co-twins. IQ accounted for nearly twice the variance in severity scores in co-twins than it did for twins with autism (28% for co-twins; 15.6% for twins with autism). For both groups, the block of brain volumes accounted for a large proportion of unique variance in severity scores (65% for twins with autism; 48% for co-twins), although only dorsolateral prefrontal volumes in autistic twins were significantly associated with severity scores ( Figure 1 ).

When measured with the Autism Diagnostic Observation Schedule, autism severity was significantly associated with neuroanatomic volumes, but not IQ scores, for twins with autism. Although IQ accounted for only 3% of the variance in autism severity (p=0.66), the combination of brain volumes accounted for 93% of the variance (p=0.005). Larger dorsal prefrontal and amygdala volumes as well as smaller vermis volumes were significant predictors ( Table 4 ). In contrast, this measure of autism severity was not significantly associated with either IQ scores (p=0.47, accounting for 38% of variance) or neuroanatomic volumes (p=0.26, accounting for 14% of variance) for the co-twins, although, again, IQ accounted for a much larger portion of variance in severity for co-twins than for autistic twins.
Discussion
Our study is the first investigation, to our knowledge, of volumetric differences of the prefrontal cortex, corpus callosum, cerebellar vermis, and mesial temporal lobe between monozygotic twins with varying levels of discordance for autism. Relative to comparison subjects, we found larger volumes of the total and right dorsolateral prefrontal cortex and smaller areas of the anterior aspect of the corpus callosum in twins with autism. Neuroanatomic structures of less affected co-twins tended to be midrange between twins with autism and typically developing comparison subjects, with the exception of vermal lobules VI and VII, which were significantly smaller in co-twins than in comparison subjects. Interestingly, the degree of neuroanatomic concordance between twin pairs varied by brain region. In addition, volumes of the dorsal prefrontal cortex, amygdala, and vermal lobules VI and VII accounted for a very large portion of the variance in scores reflecting autism severity for subjects with narrowly defined autism, whereas the same neuroanatomic structures accounted for less variance in autism severity in the co-twins.
Volumetric Differences Among Children With Autism, Co-Twins, and Comparison Subjects
Our findings of increased dorsolateral prefrontal cortical volumes in children with autism are consistent with previous observations of region-specific (as opposed to global) volumetric enlargement of the frontal lobe in young children with autism (28) . Increases in dorsolateral prefrontal tissue in children with autism may be a result of anomalies in the development of both white matter and synaptic densities (28) , which occur concurrently and in a region-specific manner (50) . In typically developing children, synapse elimination is most likely complete by age 12 in the auditory cortex, whereas synapse elimination appears to continue until mid-adolescence in the middle frontal gyrus (50) . Considering the age range of our subjects, our finding of enlarged dorsolateral prefrontal cortical volumes in the absence of hyperplasia of the total frontal lobe may be a result of the likelihood that this region is still developing and has not reached the point of substantial synapse elimination that decreases synaptic density to approximately 60% of maximum (50) .
Decreases in the genu and anterior body of the corpus callosum are consistent with previous studies that have observed reduced area and fractional anisotropy in the anterior aspects of this structure (30 , 31) . Moreover, the fibers of the anterior portion of the corpus callosum project to the prefrontal cortex (51) , suggesting that the anomalies that we observed in the dorsal prefrontal cortex may be associated with disruptions in connectivity via the corpus callosum. Diffusion tensor imaging studies are needed to specifically identify interprefrontal connectivity of the corpus callosum and the extent to which disruption in those fibers affect dorsal prefrontal morphology (52) .
Despite evidence for the amygdala’s role in social cognition and “theory of mind” (1 , 53) and studies indicating that the volume of the amygdala is larger in school-age children with autism relative to typically developing children (7 , 54) , we did not observe volumetric differences in this structure. Moreover, we did not observe an association between age and volumetric change in the amygdala, which has been reported previously. Our findings may be a result of differences in the age distribution between previous and current research samples or potentially of the effect of twinning on differences in the trajectory of amygdala development.
Lobules VI and VII of the cerebellar vermis have been the subject of numerous morphometric studies in autism since anomalies were initially observed in 1988 (12) . A recent meta-analysis (55) suggests that heterogeneity of previous findings is related to sample differences in age and IQ. Since the co-twins in our sample exhibited reductions of lobules VI and VII to a greater extent than twins with autism, who had lower IQ scores, our findings do not support the notion (55) that group differences in cerebellar vermis area decrease as age and IQ increase. Nonetheless, the moderate vermis reductions that we detected in co-twins and their more severely affected twins suggest that disruptions to the vermis may be associated with both the narrow and the broader phenotype for autism, whereas disruptions in cerebral structures may be associated with the narrow phenotype only.
Neuroanatomic Concordance Within Twin Pairs
Recent studies (36 , 56) of typically developing and adult monozygotic twins have observed a high degree of within-pair neuroanatomic concordance, with correlations of at least 0.80. In contrast, the intraclass correlation coefficients reported in the present study range from 0.65 to 0.93. The variability that we observed in the degree of neuroanatomic concordance may be a result of differences in maturational trajectories in the regions of the brain that we measured. The dorsal prefrontal cortex, amygdala, and corpus callosum mature relatively late in typically developing individuals (57) . Moreover, age-related variation in the amygdala (54) and the dorsal prefrontal cortex (2) has been noted in autism. In addition, variability in concordance may be a function of differences in plasticity of specific brain regions. For example, evidence of both experience-dependent cortical plasticity (58 , 59) and of protracted development of the prefrontal cortex (50) would suggest that in early childhood the less-affected co-twins may have had the capacity and resources to interact with environmental stimuli in a manner that supported more typical neural development of the prefrontal cortex. Future longitudinal studies of discordant monozygotic twin pairs that investigate neuroanatomic variations over time are needed to further delineate neuroanatomic growth patterns and behavioral trajectories in children with autism and their co-twins.
Association Between Volumes and Behavioral Phenotypes
Our findings are consistent with previous studies reporting associations between the prefrontal cortex, amygdala, and cerebellar vermis volumes and the core features of autism. One caveat in studies of associations between prefrontal volumes and autism features is that they vary in the specific substructures of the prefrontal cortex that are associated with social and communication deficits in children with autism (60) . In addition, both positive and negative (61) associations between social interaction/communication and the prefrontal cortex have been observed, although this difference may be a result of between-study differences in the age range of participants.
Both positive and negative associations have also been observed between amygdala volumes and comorbid/behavioral features of autism, including a positive association between amygdala volumes and the degree of anxiety (49) and social and communication deficits (1) and a negative association between amygdala volumes and social reciprocity (10) . Again, differences in the age and degree of impairment between samples may account for these inconsistencies. Taken together, however, these findings support the role that the amygdala plays in the behavioral phenotype of individuals with autism.
Associations between decreases in lobules VI and VII of the cerebellar vermis and stereotyped and restrictive behaviors have been reported in children with autism (62) and in children with fragile X syndrome, a disorder that can include core features of autism (63) . Through its connections to the brainstem and cortical areas involved in auditory processing and attention (64 – 66) , one function of the vermis is to mediate sensory stimuli (64 , 67) . Accordingly, disruptions in the connectivity of these circuits may underlie the association between decreases in this structure and severity of autistic features.
Notably, full-scale intelligence did not account for a significant portion of the variance in either of the autism severity scores for the twins with autism or their co-twins. Nonetheless, in co-twins, IQ scores accounted for a much greater proportion of variance in autism severity than in the twins with autism. These differences could be a result of the small sample size and restricted ranges of either IQ scores in the twins with autism or neuroanatomic volumes in the co-twins, all affecting power to detect associations. However, this finding may also support the notion of what Rutter and colleagues (68) referred to as a “second hit,” which leads to the development of a severe autism disorder above and beyond the risk for autism that may be conferred by either genetic vulnerability or the presence of intellectual disability (67) . Potentially, a “second hit” could consist of epigenetic and/or nonshared environmental events that moderate genetic contributions to the behavioral phenotypes of our monozygotic twins. Potentially, the monozygotic twins in our study have different DNA methylation patterns or other epigenetic differences, leading to varying environmental stimuli susceptibilities (67) and dissimilar regulation of gene expression and, ultimately, discordant behavioral phenotypes.
Limitations and Conclusions
It is important to note that the twinning process may have affected our results. To the extent that premature births or lower birth weights characterize twins to a greater extent than singletons, the neuroanatomic abnormalities observed in our autism twins may be different from those that would occur in singleton subjects with autism disorder. Accordingly, our study would be strengthened by the inclusion of monozygotic twin pairs as comparison subjects. Moreover, in order to disentangle the genetic versus shared and nonshared environmental influences that contribute to neuroanatomic abnormalities in children with autism, future studies should include both typically developing monozygotic twins as well as dizygotic twins in which at least one twin is affected with autism. The present study is further limited by the small size and large age range of the sample (which reduced power to detect other neuroanatomic differences), the utilization of different IQ measures for the twin pairs and comparison subjects, and the lack of social functioning scores for all subjects, preventing us from investigating the association between brain regions of interest and social and communication deficits across all subjects. Finally, to the extent that alterations in the frontal lobe occur during the most critical time of language and social development, the incorporation of a longitudinal design in future studies is also essential. Despite these limitations, our study replicates previous findings of an enlarged dorsolateral prefrontal cortex, reduced areas of the corpus callosum, and decreased posterior cerebellar vermis in autism and demonstrates that neuroanatomic alterations are closely associated with phenotypic features of autism in children who are severely affected.
1. Munson J, Dawson G, Abbott R, Faja S, Webb SJ, Friedman SD, Shaw D, Artru A, Dager SR: Amygdalar volume and behavioral development in autism. Arch Gen Psychiatry 2006; 63:686–693Google Scholar
2. Carper RA, Courchesne E: Localized enlargement of the frontal cortex in early autism. Biol Psychiatry 2005; 57:126–133Google Scholar
3. Hardan AY, Girgis RR, Lacerda AL, Yorbik O, Kilpatrick M, Keshavan MS, Minshew NJ: Magnetic resonance imaging study of the orbitofrontal cortex in autism. J Child Neurol 2006; 21:866–871Google Scholar
4. Kates WR, Burnette CP, Eliez S, Strunge LA, Kaplan D, Landa R, Reiss AL, Pearlson GD: Neuroanatomic variation in monozygotic twin pairs discordant for the narrow phenotype for autism. Am J Psychiatry 2004; 161:539–546Google Scholar
5. Aylward EH, Minshew NJ, Goldstein G, Honeycutt NA, Augustine AM, Yates KO, Barta PE, Pearlson GD: MRI volumes of amygdala and hippocampus in non-mentally retarded autistic adolescents and adults. Neurology 1999; 53:2145–2150Google Scholar
6. Rojas DC, Smith JA, Benkers TL, Camou SL, Reite ML, Rogers SJ: Hippocampus and amygdala volumes in parents of children with autistic disorder. Am J Psychiatry 2004; 161:2038–2044Google Scholar
7. Sparks BF, Friedman SD, Shaw DW, Aylward EH, Echelard D, Artru AA, Maravilla KR, Giedd JN, Munson J, Dawson G, Dager SR: Brain structural abnormalities in young children with autism spectrum disorder. Neurology 2002; 59:184–192Google Scholar
8. Herbert MR, Ziegler DA, Deutsch CK, O’Brien LM, Lange N, Bakardjiev A, Hodgson J, Adrien KT, Steele S, Makris N, Kennedy D, Harris GJ, Caviness VS Jr: Dissociations of cerebral cortex, subcortical and cerebral white matter volumes in autistic boys. Brain 2003; 126:1182–1192Google Scholar
9. Schumann CM, Hamstra J, Goodlin-Jones BL, Lotspeich LJ, Kwon H, Buonocore MH, Lammers CR, Reiss AL, Amaral DG: The amygdala is enlarged in children but not adolescents with autism; the hippocampus is enlarged at all ages. J Neurosci 2004; 24:6392–6401Google Scholar
10. Nacewicz BM, Dalton KM, Johnstone T, Long MT, McAuliff EM, Oakes TR, Alexander AL, Davidson RJ: Amygdala volume and nonverbal social impairment in adolescent and adult males with autism. Arch Gen Psychiatry 2006; 63:1417–1428Google Scholar
11. Zeegers M, Pol HH, Durston S, Nederveen H, Schnack H, Daalen EV, Dietz C, Engeland HV, Buitelaar J: No differences in MR-based volumetry between 2- and 7-year-old children with autism spectrum disorder and developmental delay. Brain Dev 2008; Dec 8 [Epub ahead of print]Google Scholar
12. Courchesne E, Yeung-Courchesne R, Press GA, Hesselink JR, Jernigan TL: Hypoplasia of cerebellar vermal lobules VI and VII in autism. N Engl J Med 1988; 318:1349–1354Google Scholar
13. Courchesne E, Saitoh O, Yeung-Courchesne R, Press GA, Lincoln AJ, Haas RH, Schreibman L: Abnormality of cerebellar vermian lobules VI and VII in patients with infantile autism: identification of hypoplastic and hyperplastic subgroups with MR imaging. AJR Am J Roentgenol 1994; 162:123–130Google Scholar
14. Saitoh O, Courchesne E, Egaas B, Lincoln AJ, Schreibman L: Cross-sectional area of the posterior hippocampus in autistic patients with cerebellar and corpus callosum abnormalities. Neurology 1995; 45:317–324Google Scholar
15. Hashimoto T, Tayama M, Murakawa K, Yoshimoto T, Miyazaki M, Harada M, Kuroda Y: Development of the brainstem and cerebellum in autistic patients. J Autism Dev Disord 1995; 25:1–18Google Scholar
16. Haas RH, Townsend J, Courchesne E, Lincoln AJ, Schreibman L, Yeung-Courchesne R: Neurologic abnormalities in infantile autism. J Child Neurol 1996; 11:84–92Google Scholar
17. Carper RA, Courchesne E: Inverse correlation between frontal lobe and cerebellum sizes in children with autism. Brain 2000; 123(pt 4):836–844Google Scholar
18. Hardan AY, Minshew NJ, Harenski K, Keshavan MS: Posterior fossa magnetic resonance imaging in autism. J Am Acad Child Adolesc Psychiatry 2001; 40:666–672Google Scholar
19. Courchesne E, Karns CM, Davis HR, Ziccardi R, Carper RA, Tigue ZD, Chisum HJ, Moses P, Pierce K, Lord C, Lincoln AJ, Pizzo S, Schreibman L, Haas RH, Akshoomoff NA, Courchesne RY: Unusual brain growth patterns in early life in patients with autistic disorder: an MRI study. Neurology 2001; 57:245–254Google Scholar
20. Akshoomoff N, Lord C, Lincoln AJ, Courchesne RY, Carper RA, Townsend J, Courchesne E: Outcome classification of preschool children with autism spectrum disorders using MRI brain measures. J Am Acad Child Adolesc Psychiatry 2004; 43:349–357Google Scholar
21. Webb SJ, Sparks BF, Friedman SD, Shaw DW, Giedd J, Dawson G, Dager SR: Cerebellar vermal volumes and behavioral correlates in children with autism spectrum disorder. Psychiatry Res 2009; 172:61–67Google Scholar
22. Courchesne E, Press GA, Yeung-Courchesne R: Parietal lobe abnormalities detected with MR in patients with infantile autism. AJR Am J Roentgenol 1993; 160:387–393Google Scholar
23. Egaas B, Courchesne E, Saitoh O: Reduced size of corpus callosum in autism. Arch Neurol 1995; 52:794–801Google Scholar
24. Manes F, Piven J, Vrancic D, Nanclares V, Plebst C, Starkstein SE: An MRI study of the corpus callosum and cerebellum in mentally retarded autistic individuals. J Neuropsychiatry Clin Neurosci 1999; 11:470–474Google Scholar
25. Hardan AY, Minshew NJ, Keshavan MS: Corpus callosum size in autism. Neurology 2000; 55:1033–1036Google Scholar
26. Vidal CN, Nicolson R, DeVito TJ, Hayashi KM, Geaga JA, Drost DJ, Williamson PC, Rajakumar N, Sui Y, Dutton RA, Toga AW, Thompson PM: Mapping corpus callosum deficits in autism: an index of aberrant cortical connectivity. Biol Psychiatry 2006; 60:218–225Google Scholar
27. Boger-Megiddo I, Shaw DW, Friedman SD, Sparks BF, Artru AA, Giedd JN, Dawson G, Dager SR: Corpus callosum morphometrics in young children with autism spectrum disorder. J Autism Dev Disord 2006; 36:733–739Google Scholar
28. Carper RA, Moses P, Tigue ZD, Courchesne E: Cerebral lobes in autism: early hyperplasia and abnormal age effects. Neuroimage 2002; 16:1038–1051Google Scholar
29. Courchesne E, Pierce K: Brain overgrowth in autism during a critical time in development: implications for frontal pyramidal neuron and interneuron development and connectivity. Int J Dev Neurosci 2005; 23:153–170Google Scholar
30. Barnea-Goraly N, Kwon H, Menon V, Eliez S, Lotspeich L, Reiss AL: White matter structure in autism: preliminary evidence from diffusion tensor imaging. Biol Psychiatry 2004; 55:323–326Google Scholar
31. Keller TA, Kana RK, Just MA: A developmental study of the structural integrity of white matter in autism. Neuroreport 2007; 18:23–27Google Scholar
32. Howard MA, Cowell PE, Boucher J, Broks P, Mayes A, Farrant A, Roberts N: Convergent neuroanatomical and behavioural evidence of an amygdala hypothesis of autism. Neuroreport 2000; 11:2931–2935Google Scholar
33. Haznedar MM, Buchsbaum MS, Wei TC, Hof PR, Cartwright C, Bienstock CA, Hollander E: Limbic circuitry in patients with autism spectrum disorders studied with positron emission tomography and magnetic resonance imaging. Am J Psychiatry 2000; 157:1994–2001Google Scholar
34. Lenroot RK, Giedd JN: Brain development in children and adolescents: insights from anatomical magnetic resonance imaging. Neurosci Biobehav Rev 2006; 30:718–729Google Scholar
35. Giedd JN, Schmitt JE, Neale MC: Structural brain magnetic resonance imaging of pediatric twins. Hum Brain Mapp 2007; 28:474–481Google Scholar
36. Wallace GL, Eric Schmitt J, Lenroot R, Viding E, Ordaz S, Rosenthal MA, Molloy EA, Clasen LS, Kendler KS, Neale MC, Giedd JN: A pediatric twin study of brain morphometry. J Child Psychol Psychiatry 2006; 47:987–993Google Scholar
37. Lord C, Rutter M, Le Couteur A: Autism Diagnostic Interview—Revised: a revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J Autism Dev Disord 1994; 24:659–685Google Scholar
38. Lord C, Risi S, Lambrecht L, Cook EH Jr, Leventhal BL, DiLavore PC, Pickles A, Rutter M: The Autism Diagnostic Observation Schedule—Generic: a standard measure of social and communication deficits associated with the spectrum of autism. J Autism Dev Disord 2000; 30:205–223Google Scholar
39. Achenbach TM, Edelbrock C: Manual for the Child Behavior Checklist and Revised Child Behavior Profile. Burlington, Vt, Queen City Printers, 1983Google Scholar
40. Subramaniam B, Hennessey J, Rubin M, Beach L, Reiss A: Software and methods for quantitative imaging in neuroscience, in Neuroinformatics: An Overview of the Human Brain Project. Edited by Koslow S, Huerta M. Hillsdale, NJ, Lawrence Erlbaum Associates, 1997, pp 335–360Google Scholar
41. Baumgardner TL, Singer HS, Denckla MB, Rubin MA, Abrams MT, Colli MJ, Reiss AL: Corpus callosum morphology in children with Tourette syndrome and attention deficit hyperactivity disorder. Neurology 1996; 47:477–482Google Scholar
42. Reiss AL, Aylward E, Freund LS, Joshi PK, Bryan RN: Neuroanatomy of fragile X syndrome: the posterior fossa. Ann Neurol 1991; 29:26–32Google Scholar
43. Kates WR, Abrams MT, Kaufmann WE, Breiter SN, Reiss AL: Reliability and validity of MRI measurement of the amygdala and hippocampus in children with fragile X syndrome. Psychiatry Res 1997; 75:31–48Google Scholar
44. Gur RE, Cowell PE, Latshaw A, Turetsky BI, Grossman RI, Arnold SE, Bilker WB, Gur RC: Reduced dorsal and orbital prefrontal gray matter volumes in schizophrenia. Arch Gen Psychiatry 2000; 57:761–768Google Scholar
45. Aylward EH, Augustine A, Li Q, Barta PE, Pearlson GD: Measurement of frontal lobe volume on magnetic resonance imaging scans. Psychiatry Res 1997; 75:23–30Google Scholar
46. Thorndike RL, Hagen EP, Sattler JM: Stanford-Binet Intelligence Scale: Guide for Administering and Scoring, 4th ed. Chicago, Riverside Publishing, 1986Google Scholar
47. Wechsler D: Manual for the Wechsler Intelligence Scale for Children, 3rd ed. San Antonio, Tex, Psychological Corp, 1991Google Scholar
48. Hollingshead AB, Redlich FC: Social Class and Mental Illness. New York, John Wiley and Sons, 1958Google Scholar
49. Juranek J, Filipek PA, Berenji GR, Modahl C, Osann K, Spence MA: Association between amygdala volume and anxiety level: magnetic resonance imaging (MRI) study in autistic children. J Child Neurol 2006; 21:1051–1058Google Scholar
50. Huttenlocher PR, Dabholkar AS: Regional differences in synaptogenesis in human cerebral cortex. J Comp Neurol 1997; 387:167–178Google Scholar
51. Hofer S, Frahm J: Topography of the human corpus callosum revisited: comprehensive fiber tractography using diffusion tensor magnetic resonance imaging. Neuroimage 2006; 32:989–994Google Scholar
52. Miyata J, Hirao K, Namiki C, Fukuyama H, Okada T, Miki Y, Hayashi T, Murai T: Interfrontal commissural abnormality in schizophrenia: tractography-assisted callosal parcellation. Schizophr Res 2007; 97:236–241Google Scholar
53. Shaw P, Lawrence EJ, Radbourne C, Bramham J, Polkey CE, David AS: The impact of early and late damage to the human amygdala on “theory of mind” reasoning. Brain 2004; 127:1535–1548Google Scholar
54. Schumann CM, Amaral DG: Stereological analysis of amygdala neuron number in autism. J Neurosci 2006; 26:7674–7679Google Scholar
55. Stanfield AC, McIntosh AM, Spencer MD, Philip R, Gaur S, Lawrie SM: Towards a neuroanatomy of autism: a systematic review and meta-analysis of structural magnetic resonance imaging studies. Eur Psychiatry 2008; 23:289–299Google Scholar
56. White T, Andreasen NC, Nopoulos P: Brain volumes and surface morphology in monozygotic twins. Cereb Cortex 2002; 12:486–493Google Scholar
57. Lenroot RK, Giedd JN: The changing impact of genes and environment on brain development during childhood and adolescence: initial findings from a neuroimaging study of pediatric twins. Dev Psychopathol 2008; 20:1161–1175Google Scholar
58. Klintsova AY, Greenough WT: Synaptic plasticity in cortical systems. Curr Opin Neurobiol 1999; 9:203–208Google Scholar
59. Markham JA, Greenough WT: Experience-driven brain plasticity: beyond the synapse. Neuron Glia Biol 2004; 1:351–363Google Scholar
60. Kennedy DP, Redcay E, Courchesne E: Failing to deactivate: resting functional abnormalities in autism. Proc Natl Acad Sci U S A 2006; 103:8275–8280Google Scholar
61. Girgis RR, Minshew NJ, Melhem NM, Nutche JJ, Keshavan MS, Hardan AY: Volumetric alterations of the orbitofrontal cortex in autism. Prog Neuropsychopharmacol Biol Psychiatry 2007; 31:41–45Google Scholar
62. Pierce K, Courchesne E: Evidence for a cerebellar role in reduced exploration and stereotyped behavior in autism. Biol Psychiatry 2001; 49:655–664Google Scholar
63. Mazzocco MM, Kates WR, Baumgardner TL, Freund LS, Reiss AL: Autistic behaviors among girls with fragile X syndrome. J Autism Dev Disord 1997; 27:415–435Google Scholar
64. Crispino L, Bullock TH: Cerebellum mediates modality-specific modulation of sensory responses of midbrain and forebrain in rat. Proc Natl Acad Sci U S A 1984; 81:2917–2920Google Scholar
65. Joseph JW, Shambes GM, Gibson JM, Welker W: Tactile projections to granule cells in caudal vermis of the rat’s cerebellum. Brain Behav Evol 1978; 15:141–149Google Scholar
66. Tang ZW, Zhang SQ: The cerebellar projection from the reticular formation of the brain stem in the rabbit: an experimental study using HRP as a retrograde tracer. Anat Embryol (Berl) 1987; 175:521–526Google Scholar
67. Schanen NC: Epigenetics of autism spectrum disorders. Hum Mol Genet 2006; 15(spec no 2):R138–R150Google Scholar
68. Rutter M, Bailey A, Simonoff E, Pickles A: Genetic influences and autism, in Handbook of Autism and Pervasive Developmental Disorder. Edited by Cohen D, Volkmar F. New York, John Wiley and Sons, 1997, pp 370–387Google Scholar

