
Low Extraversion and High Neuroticism as Indices of Genetic and Environmental Risk for Social Phobia, Agoraphobia, and Animal Phobia
Abstract
Objective: The authors examined the extent to which two major personality dimensions (extraversion and neuroticism) index the genetic and environmental risk for three phobias (social phobia, agoraphobia, and animal phobia) in twins ascertained from a large, population-based registry. Method: Lifetime phobias and personality traits were assessed through diagnostic interview and self-report questionnaire, respectively, in 7,800 twins from female-female, male-male, and opposite-sex pairs. Sex-limited trivariate Cholesky structural equation models were used to decompose the correlations among extraversion, neuroticism, and each phobia. Results: In the best-fitting models, genetic correlations were moderate and negative between extraversion and both social phobia and agoraphobia, and that between extraversion and animal phobia was effectively zero. Genetic correlations were high and positive between neuroticism and both social phobia and agoraphobia, and that between neuroticism and animal phobia was moderate. All of the genetic risk factors for social phobia and agoraphobia were shared with those that influence extraversion and neuroticism; in contrast, only a small proportion of the genetic risk factors for animal phobia (16%) was shared with those that influence personality. Shared environmental experiences were not a source of correlations between personality traits and phobias, and unique environmental correlations were relatively modest. Conclusion: Genetic factors that influence individual variation in extraversion and neuroticism appear to account entirely for the genetic liability to social phobia and agoraphobia, but not animal phobia. These findings underline the importance of both introversion (low extraversion) and neuroticism in some psychiatric disorders.
Phobias run in families, and twin studies suggest that the modest familial aggregation observed in general population samples is mainly due to genetic factors (1) . Exactly what is inherited, though, is unclear. In particular, it is of interest to determine to what extent the familial vulnerability to phobias is indexed by basic personality traits, such as extraversion and neuroticism (2 , 3) . Low extraversion (introversion) and high neuroticism enhance aversive conditioning in laboratory settings (4 – 6) .
Extraversion and neuroticism are found in almost all personality nosologies (7) . Extraversion refers to a person’s tendency to be venturesome, energetic, assertive, and sociable and to experience positive emotions (e.g., joy). Eysenck theorized that introverts have higher levels of activity in the ascending reticular activating system and are more “aroused” than extraverts, in that introverts are more distractible in high-stimulus environments and perform better at prolonged, monotonous tasks (4) . Though the physical basis of extraversion remains under investigation (6 , 8) , there is some empirical support for Eysenck’s theory (4 , 9) . Neuroticism refers to a person’s general tendency to experience negative emotions (e.g., nervousness, sadness, and anger). Eysenck theorized that neuroticism reflects a person’s characteristic limbic “excitability,” based on autonomic activation patterns (4) .
The relationship between neuroticism and anxiety/depressive disorders is widely recognized; less well known is the finding that introversion is also consistently associated with some of these conditions. Social phobia and agoraphobia have particularly strong associations with both introversion and neuroticism. In contrast, relationships between these traits and specific phobias (e.g., animal phobia) tend to be relatively weak (10 , 11) .
Extraversion and neuroticism are moderately heritable (12) and may contribute part of the heritable basis of phobias. A number of family studies have partially and indirectly addressed this hypothesis for social phobia. For example, trait anxiety, harm avoidance, and behavioral inhibition all combine aspects of introversion and neuroticism (5 , 13 , 14) and appear familially related to social phobia (15 , 16) . To our knowledge, familial relationships between personality traits and agoraphobia have received less attention; however, agoraphobia appears familially related to behavioral inhibition (17) . In contrast, behavioral inhibition does not appear familially related to specific phobias (16) ; this finding is consistent with weak phenotypic relationships between personality traits and specific phobias.
Thus, extant family studies suggest that introversion and neuroticism may index the heritable liability to social phobia and agoraphobia, but they do not address whether the familial relationships are due to genetic or shared environmental factors (e.g., children could “learn” shy behavior from their social phobic or agoraphobic parents). Furthermore, none of the studies have tested whether there are independent contributions of introversion and neuroticism, since they all used measures that combine the two dimensions. Finally, to our knowledge, no studies have assessed whether there are inherited characteristics that are specific to phobias, beyond those indexed by personality traits.
In the current study, we employed large twin samples to address these issues for social phobia, agoraphobia, and a common specific phobia, animal phobia. On the basis of extant phenotypic and family studies, we predicted that both introversion and neuroticism would substantially index the genetic vulnerability to social phobia and agoraphobia. Also on the basis of extant studies, we expected that the results for animal phobia would contrast with those for social phobia and agoraphobia, in having weaker phenotypic and genetic relationships to personality traits.
Method
Subjects
The twin data in this report derive from two interrelated projects involving participants in the population-based Virginia Twin Registry, details of which are described elsewhere (18 , 19) . Briefly, twins from female-female pairs were eligible if they were white and born between 1934 and 1971, and twins from male-male and male-female pairs were eligible if they were white and born between 1940 and 1974. The current study utilizes data from the first interview wave of female-female twins and the second interview wave of male-male and male-female twins. In the interviews of female-female twins, 2,163 subjects were interviewed, including 1,033 complete pairs, 58% of which were monozygotic and 42% dizygotic; 89% of the subjects were interviewed in person and 11% by telephone. In the interviews of male-male twins, 2,939 subjects were interviewed, including 1,199 complete pairs, 59% of which were monozygotic and 41% dizygotic. In the interviews of opposite-sex twins, 2,698 subjects were interviewed, including 1,070 complete pairs. Of the male-male and male-female twins, 80% were interviewed in person and 20% by telephone. Zygosity determinations using standard questions and photographs were validated by using genetic marker data, with an error rate of less than 5% (19) . The interviews at these waves utilized identical phobia screening questions, and these particular waves were the first time in either study in which the participants were assessed for phobias. The assessments were also comparable in that they employed identical self-administered personality questionnaires. The average age at interview was 30.1 years (SD=7.6, range=17–55) in the female-female sample and 37.0 years (SD=9.1, range=20–58) in the sample of male-male and male-female twins. Written informed consent was obtained before in-person interviews, and verbal consent was obtained before telephone interviews. The projects were approved by the Committee for the Conduct of Human Research at Virginia Commonwealth University.
Measures
Lifetime phobias were assessed with an adaptation of the Diagnostic Interview Schedule version III-A (20) . The assessed social fears included meeting new people, giving a speech, using public bathrooms, and eating in public; the assessed agoraphobia-related fears included going out of the house alone, being in crowds, and being in open spaces; and the assessed animal-related fears included spiders, “bugs,” mice, snakes, bats, and other animals. In this study, a phobia was diagnosed if interviewers judged that the particular fear and related avoidance interfered significantly in a respondent’s life. The interviewers were carefully trained and supervised mental health workers with at least a master’s degree or a bachelor’s degree and 2 years of clinical experience. Two senior staff members reviewed each interview for completeness and consistency. The members of each twin pair were assessed by different interviewers who were blind to clinical information about the co-twins. All data were entered twice to minimize data-entry errors.
Values indicating test-retest reliability for social phobia, agoraphobia, and animal phobia, based on the current methods, were in the modest to moderate range (kappas for females: 0.47, 0.52, 0.49, respectively; tetrachoric correlations for females: 0.79, 0.84, 0.78; kappas for males: 0.37, 0.27, 0.17; correlations for males: 0.72, 0.76, 0.43) (21 , 22) ; reliability was unrelated to zygosity. The lifetime prevalences of social phobia, agoraphobia, and animal phobia in females were 14.1%, 9.1%, and 10.5%, respectively; in males these were 6.3%, 4.0%, and 5.2%, respectively (22 , 23) .
Extraversion was assessed with eight items and neuroticism was evaluated with 12 items from the short form of the self-administered Eysenck Personality Questionnaire (24) . For extraversion, the Cronbach alpha value was 0.83 for the female-female sample and 0.82 for the sample with male-male and male-female twins; the comparable estimates for neuroticism were 0.84 and 0.85, respectively. Internal consistency was unrelated to zygosity. Personality scores were analyzed as ordinal variables (extraversion range=0–8, neuroticism range=0–12).
Statistical Analysis
We used SAS version 9.1 (SAS Institute, Cary, N.C.) to calculate polychoric and tetrachoric correlations and SPSS version 12.0 (SPSS, Chicago) in logistic regression analyses. The purpose of including subjects from the comparable female-female and male-male/male-female projects was to maximize sample size and statistical power. Since it would not be justified to assume genetic homogeneity across sexes, we explicitly tested for global sex differences prior to testing the hypotheses of interest for this study. Specifically, we applied sex-limited trivariate Cholesky structural equation models to the twin data; these allowed us to assess genetic and environmental liabilities shared among extraversion, neuroticism, and each phobia, taking possible sex differences into account (25) . The sex-limitation model allowed for two types of possible sex differences: 1) sex-specific genetic effects (which would imply some nonoverlap of the genes that influence the phenotypes in men and women) and 2) differences in the magnitude of effects of the same underlying latent genetic and environmental factors (quantitative sex differences). The models imposed a stratified structure on the latent factors hypothesized to determine the measured phenotypes, with the first group of factors—additive genetic (A1), female-specific additive genetic (A fs 1), shared common environmental (C1), and unique environmental (E1)—influencing extraversion, neuroticism, and the phobia; the second group of factors—A2, A fs 2, C2, and E2—influencing only neuroticism and the phobia; and the third group—A3, A fs 3, C3, and E3—influencing only the phobia. The specific ordering of extraversion and neuroticism was arbitrary and not of particular interest; we chose to include the phobias last since we were particularly interested in whether or not there were genetic influences that were unique to each phobia, not shared with those that influence extraversion or neuroticism. The model is illustrated in simplified form in the top part of Figure 1 .
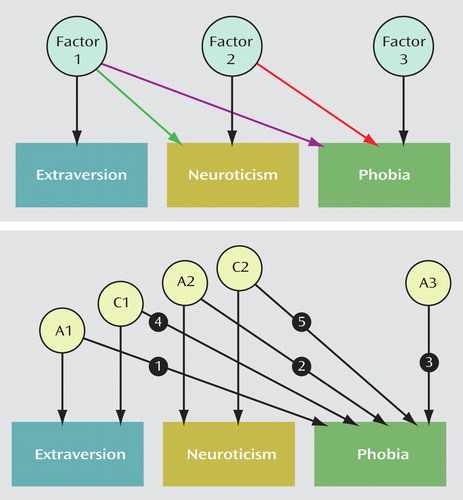
a The top panel represents one twin and a single generic set of “factors,” though the full model included three sets for males and four sets for females, i.e., additive genetic (A), female-specific additive genetic, shared (or common) environmental (C), and unique environmental. Latent variables appear in circles, while observed (measured) variables appear in rectangles. The bottom panel shows particular factors and paths of interest for hypothesis testing, referred to in the Results section. Path 1 represents additive genetic effects that influence both extraversion and phobia; path 2 represents additive genetic effects that influence both neuroticism and phobia; path 3 represents additive genetic effects that influence only phobia; path 4 represents shared environmental effects that influence both extraversion and phobia; and path 5 represents shared environmental effects that influence both neuroticism and phobia.
We fit models to the raw data using the Mx program (26) . Model testing began with each full model, including all of the aforementioned sources of variance. Thresholds for personality traits and phobias were allowed to differ between males and females, given sex differences in means and prevalences, respectively. Model parameters and indices that characterized the fit of each full model were calculated, and then the full models were compared with nested submodels created by eliminating or constraining parameters in a stepwise fashion. The goal was to identify the most parsimonious model that sufficiently described the data. The fit of nested submodels was compared by taking the difference between negative two times the log likelihood of the data (–2LL) for each full model and respective submodels; under certain regularity conditions, these differences follow a chi-square distribution, with degrees of freedom equal to the difference in degrees of freedom between the two models. More parsimonious models (i.e., those with fewer parameters) are considered preferable if they do not provide a significantly worse fit to the data. To operationalize this, we used the Akaike information criterion (AIC) statistic, calculated as the model chi-square value minus two times the degrees of freedom. Lower AIC values suggest a better balance of explanatory power and complexity.
We modeled each phobia separately. Prior to hypothesis testing, we created simplified sex-limited “baseline” models. In a stepwise fashion, we 1) eliminated the six female-specific additive genetic parameters, 2) equated the six nonstandardized additive genetic parameters across sexes, and 3) equated the six nonstandardized shared environmental parameters across sexes. In models for each phobia, each of these changes resulted in a favorable balance of model fit and parsimony (lower AIC). However, for each phobia, when we also attempted to equate the six unique environmental paths across sexes, there was a significant loss of fit. Equating unique environmental paths across sexes forced the proportion of genetic variance to be identical across sexes, although in this study, cross-twin, within-trait correlations for female monozygotic twins were larger than the corresponding correlations for male monozygotic twins for all phenotypes except animal phobia (see table in online data supplement, correlations in blue). That is, there was evidence for global quantitative sex differences (the same genes having larger effects in women). We accounted for this by allowing separate unique environmental parameters for each sex in our hypothesis-testing models.
Results
Table 1 shows polychoric and tetrachoric correlations for extraversion, neuroticism, and phobias in monozygotic and dizygotic twins. First, note the substantial negative within-person correlation between extraversion and social phobia and the positive within-person correlation between neuroticism and social phobia. These phenotypic correlations are consistent with prior observations; i.e., persons with social phobia are typically low in extraversion and/or high in neuroticism. Next, note that cross-twin correlations between extraversion and social phobia and between neuroticism and social phobia are larger in absolute value in monozygotic versus dizygotic twins; this suggests that genetic factors that affect extraversion and those that affect neuroticism also affect social phobia.
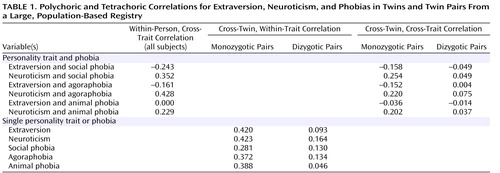
For agoraphobia, the pattern is similar to that for social phobia. In contrast, animal phobia was not at all phenotypically related to extraversion, and it was relatively weakly related to neuroticism. The genetic factors that influence animal phobia appear to overlap with those that influence neuroticism.
Table 2 shows the results of our model-fitting procedures (again, separate for each phobia). Model 1, our “baseline” model, includes the sex limitation of the unique environmental parameters specified earlier. In model 2, we set the additive genetic paths between extraversion and each phobia (path 1 in the lower part of Figure 1 ) to zero. For the social phobia and agoraphobia models, this change resulted in a substantial loss of fit (higher AIC values), indicating that extraversion and these phobias share a portion of genetic determinants. In contrast, the reduced model fit the animal phobia data well, suggesting that extraversion and animal phobia are genetically unrelated. In model 3, we set the additive genetic paths between neuroticism and each phobia (path 2 in Figure 1 ) to zero, and there was substantial worsening of fit in every case. This indicates that neuroticism and all three phobias share a significant portion of genetic determinants, separate from those of extraversion. In model 4, we dropped the phobia-specific additive genetic paths (path 3 in Figure 1 ), with a substantial improvement in fit for social phobia and agoraphobia but a worsening of fit for animal phobia. This suggests that the quantitative measures extraversion and neuroticism themselves fully index the genetic liability to social phobia and agoraphobia, but not animal phobia (i.e., there was evidence for substantial genetic factors specific to animal phobia). In model 5, we set the common environmental paths between extraversion and each phobia and between neuroticism and each phobia (paths 4 and 5 in Figure 1 ) to zero. In each case, these changes produced further improvements in the AIC (lower values), suggesting that environmental experiences shared by twins contribute little to the covariation between these personality traits and phobias.
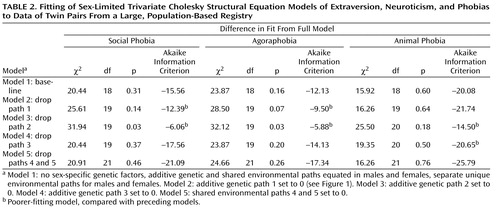
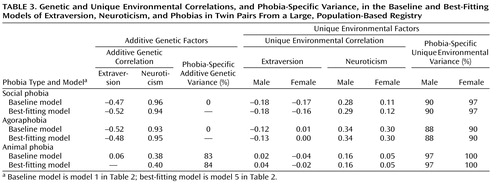
Table 3 shows results for each baseline model (model 1) and best-fitting model (model 5) in terms of additive genetic correlations (r g values—estimates of the degree to which the same genetic factors influence two variables) and unique environmental correlations (r e values—estimates of the degree to which the same environmental factors influence two variables), as well as the percentage of phobia-specific additive genetic and unique environmental variance, i.e., that not shared with extraversion or neuroticism. Shared environmental parameters are not shown, since the corresponding path coefficients were quite small. The baseline and best-fitting models were very similar. The estimates of r g were substantial for the additive genetic correlations between extraversion and social phobia or agoraphobia (negative correlations) and between neuroticism and these phobias (positive correlations), while the r e estimates were relatively small. The corresponding r g values for animal phobia were considerably smaller than those for social phobia and agoraphobia. The estimate of social phobia-specific and agoraphobia-specific genetic variance was 0%, while the estimate of animal phobia-specific genetic variance was greater than 80%.
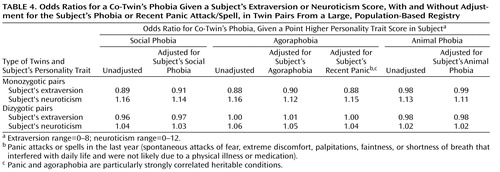
Since having a phobia or recent panic might affect personality measures (27) , we conducted logistic regression analyses to determine the extent to which a subject’s phobia or panic could confound the relationship between the subject’s personality traits and her or his co-twin’s phobia. As shown in Table 4 , accounting for a subject’s phobia or panic had little effect on these relationships (the odds ratios were comparable when a subject’s phobia or panic was taken into account). Note that these analyses likely overcontrol for psychopathology, since high neuroticism and behavioral inhibition appear to predict later onset of panic, social phobia, and perhaps agoraphobia (14 , 28 , 29) .
Discussion
Our results suggest that the familial co-occurrence of certain personality traits and phobias has a genetic, not a shared environmental, basis. Further, genetic factors that influence individual variation in extraversion and neuroticism appear to account entirely for the genetic liability to social phobia and agoraphobia, but not animal phobia. Finally, our results indicate the importance of both introversion and neuroticism as personality endophenotypes for social phobia and agoraphobia. Though geneticists often seek a single basic dimension for an endophenotype, our results suggest that the greatest genetic risk for social phobia or agoraphobia involves genetic liability to both low extraversion and high neuroticism.
The extent to which introversion and neuroticism index the genetic vulnerability to social phobia and agoraphobia here is particularly noteworthy (estimated at 100%). For comparison, when we used this cohort and similar methods, estimates of the extent to which neuroticism indexes the genetic vulnerability to major depressive disorder, generalized anxiety disorder, and panic disorder were 36% (r g =0.60), 59% (r g =0.77), and 48% (r g =0.69), respectively (30) ; none of these conditions was associated with introversion (31) . As expected, personality traits indexed only a small portion of the heritable basis of animal phobia in this study. The heritable basis of animal phobia may involve “preparedness” for aversive conditioning to specific stimuli (32) , which neither affects nor is affected by personality to a substantial degree.
In a previous multivariate twin study of neuroticism and internalizing disorders, we reported results for two broad genetic factors, one that influenced neuroticism and each disorder and a second, independent of neuroticism, that influenced major depression, generalized anxiety disorder, and panic disorder (the phobias did not have substantial loadings on this second factor) (30) . The current study suggests that if a third genetic factor linked to extraversion were added to the earlier model, this third factor would account for the remaining genetic variance of social phobia and agoraphobia, though not animal phobia.
To our knowledge, this is the first study to demonstrate genetic overlap between introversion and psychiatric disorders. Notably, the phenotypic and genetic correlations between extraversion and social phobia or agoraphobia were smaller in absolute value than those for neuroticism here. This contrasts with results from another general population study (11) in which we used a different personality measure (33) and psychiatrist diagnoses; in that study, these phobias were more strongly related to introversion than to neuroticism. Thus, had we used an alternative measure of extraversion in this study (e.g., one that explicitly includes positive emotionality), the genetic correlations with social phobia or agoraphobia might have been larger in absolute value.
Some have argued that combining introversion with neuroticism, as in trait anxiety (5) or harm avoidance (13) , provides a more parsimonious construct to describe the presumed temperamental vulnerability to anxiety and depressive disorders. However, since introversion is not consistently associated with all of these disorders (10 , 11 , 31) , it seems useful to consider extraversion and neuroticism separately. It remains an open question whether the physical bases of neuroticism and introversion are most usefully construed as constituting a single neurobiological system (5 , 13) or two interacting systems (4) .
Our results have obvious relevance for molecular genetics. As in other anxiety and depressive disorders, finding genes that influence neuroticism should be valuable in determining the etiology of phobias. Several groups are currently searching the genome for loci that influence neuroticism (for instance, see references 34 and 35 ). Our results suggest that determining the genetic basis of introversion/extraversion should also be valuable in determining the etiologies of social phobia and agoraphobia. We know of no current systematic studies to identify genetic loci that influence introversion/extraversion, though candidate gene studies exist for this phenotype (for instance, see reference 36 ). We hope that our findings stimulate further genetic research on extraversion.
While considering the implications of our study for clinical practice, prevention, and research, the limitations of our cross-sectional method should be borne in mind. That is, though our statistical model specifies that latent factors affect all of the phenotypes of interest directly (i.e., there are no causal arrows between the measured variables), it is conceivable that this is not the case. For example, it is possible that genetic and unique environmental factors affect personality traits directly and that introversion and/or neuroticism are themselves true risk factors for social phobia or agoraphobia. This would be consistent with theories regarding the effects of introversion and neuroticism on aversive conditioning (e.g., in the context of social evaluation; noisy, close, or exposed environments; and/or anxiety or panic symptoms) (4 , 5) and with theories that relate extraversion to reward-seeking behavior (5) (i.e., extraverts should find venturing into unfamiliar or bustling public environments pleasurable). However, the hypothesis that personality traits mediate genetic risk for phobias and alternatives (e.g., personality traits are simply markers of genetic risk for social phobia or agoraphobia) are difficult to test with cross-sectional data when patterns of inheritance are similar across phenotypes, as in the current study (25) ; a longitudinal study would be more appropriate (for instance, see reference 37 ). The results in Table 4 suggest that “scar” and/or state effects could not account for much of the observed covariance in this study; nevertheless, a conservative conclusion is that low extraversion and high neuroticism are powerful indices of genetic risk for social phobia and agoraphobia in adults .
Second, our models require several assumptions (25) , including the absence of assortative mating (likely minimal for the phenotypes considered here [ 12 , 38 ]) and the independence and additivity of the latent variables. Gene-environment interaction could affect twin similarity in either direction, depending on whether both twins are exposed to the specific environmental factor in question; to our knowledge, gene-environment interactions and correlations have yet to be demonstrated for the phenotypes studied here. It is important that the assumption of equal relevant shared environmental experiences for monozygotic and dizygotic twins appears valid here (22 , 23 , 39) .
Third, though nonadditive genetic effects have been detected for personality traits (12) , we did not model these here. Given the inclusion of binary phenotypes (phobias), we had inadequate power to discriminate nonadditive from additive genetic effects.
Fourth, unique environmental effects and measurement error are confounded in our models; this may bias our unique environmental correlation estimates downward. Nevertheless, our low unique environmental correlations for personality traits and phobias parallel the low unique environmental correlation for avoidant personality traits and social phobia in the study by Reichborn-Kjennerud et al. that appears elsewhere in this issue of the Journal. In both studies, unique environmental experiences mainly seem to account for phenotypic differences in persons with similar genetic vulnerabilities.
Fifth, our samples were made up entirely of Caucasian twins born in Virginia. Thus, our results may not generalize to individuals from other backgrounds.
1. Hettema JM, Neale MC, Kendler KS: A review and meta-analysis of the genetic epidemiology of anxiety disorders. Am J Psychiatry 2001; 158:1568–1578Google Scholar
2. Smoller JW, Tsuang MT: Panic and phobic anxiety: defining phenotypes for genetic science. Am J Psychiatry 1998; 155:1152–1162Google Scholar
3. Andrews G, Stewart G, Allen R, Henderson AS: The genetics of six neurotic disorders: a twin study. J Affect Disord 1990; 19:23–29Google Scholar
4. Eysenck HJ, Eysenck MW: Personality and Individual Differences: A Natural Science Approach. New York, Plenum, 1985, pp 191–309Google Scholar
5. Gray JA, McNaughton N: Putting Humpty Dumpty together again: the anxious personality and its inheritance, in The Neuropsychology of Anxiety, 2nd ed. New York, Oxford University Press, 2000, pp 333–349Google Scholar
6. Rauch SL, Milad MR, Orr SP, Quinn BT, Fischl B, Pitman RK: Orbitofrontal thickness, retention of fear extinction, and extraversion. Neuroreport 2005; 16:1909–1912Google Scholar
7. Watson D, Clark LA, Harkness AR: Structures of personality and their relevance to psychopathology. J Abnorm Psychol 1994; 103:18–31Google Scholar
8. Canli T (ed): Biology of Personality and Individual Differences. New York, Guilford, 2006Google Scholar
9. Johnson DL, Wiebe JS, Gold SM, Andreasen NC, Hichwa RD, Watkins GL, Boles Ponto LL: Cerebral blood flow and personality: a positron emission tomography study. Am J Psychiatry 1999; 156:252–257Google Scholar
10. Bienvenu OJ, Stein MB: Personality and anxiety disorders: a review. J Personal Disord 2003; 17:139–151Google Scholar
11. Bienvenu OJ, Samuels JF, Costa PT, Reti IM, Eaton WW, Nestadt G: Anxiety and depressive disorders and the five-factor model of personality: a higher- and lower-order personality trait investigation in a community sample. Depress Anxiety 2004; 20:92–97Google Scholar
12. Eaves LJ, Heath AC, Martin NG, Maes H, Neale MC, Kendler KS, Kirk K, Corey L: Comparing the biological and cultural inheritance of personality and social attitudes in the Virginia 30,000 study of twins and their relatives. Twin Res 1999; 2:62–80Google Scholar
13. Cloninger CR: A unified biosocial theory of personality and its role in the development of anxiety states. Psychiatr Dev 1986; 3:167–226Google Scholar
14. Hayward C, Killen JD, Kraemer HC, Taylor CB: Linking self-reported childhood behavioral inhibition to adolescent social phobia. J Am Acad Child Adolesc Psychiatry 1998; 37:1308–1316Google Scholar
15. Stein MB, Chartier MJ, Lizak MV, Jang KJ: Familial aggregation of anxiety-related quantitative traits in generalized social phobia: clues to understanding “disorder” heritability? Am J Med Genet 2001; 105:79–83Google Scholar
16. Rosenbaum JF, Biederman J, Hirshfeld DR, Bolduc EA, Faraone SV, Kagan J, Snidman N, Reznick JS: Further evidence of an association between behavioral inhibition and anxiety disorders: results from a family study of children from a non-clinical sample. J Psychiatr Res 1991; 25:49–65Google Scholar
17. Rosenbaum JF, Biederman J, Gersten M, Hirshfeld DR, Meminger SR, Herman JB, Kagan J, Reznick JS, Snidman N: Behavioral inhibition in children of parents with panic disorder and agoraphobia: a controlled study. Arch Gen Psychiatry 1988; 45:463–470Google Scholar
18. Kendler KS, Neale MC, Kessler RC, Heath AC, Eaves LJ: A population-based twin study of major depression in women: the impact of varying definitions of illness. Arch Gen Psychiatry 1992; 49:257–266Google Scholar
19. Kendler KS, Prescott CA: A population-based twin study of lifetime major depression in men and women. Arch Gen Psychiatry 1999; 56:39–44Google Scholar
20. Robins LN, Helzer JE: Diagnostic Interview Schedule (DIS), version III-A. St Louis, Washington University School of Medicine, 1985Google Scholar
21. Kendler KS, Karkowski LM, Prescott CA: Fears and phobias: reliability and heritability. Psychol Med 1999; 29:539–553Google Scholar
22. Kendler KS, Myers J, Prescott CA, Neale MC: The genetic epidemiology of irrational fears and phobias in men. Arch Gen Psychiatry 2001; 58:257–265Google Scholar
23. Kendler KS, Neale MC, Kessler RC, Heath AC, Eaves LJ: The genetic epidemiology of phobias in women: the interrelationship of agoraphobia, social phobia, situational phobia, and simple phobia. Arch Gen Psychiatry 1992; 49:273–281Google Scholar
24. Eysenck SBG, Eysenck HJ, Barrett P: A revised version of the psychoticism scale. Pers Individ Dif 1985; 6:21–29Google Scholar
25. Neale MC, Cardon LR: Methodology for Genetic Studies of Twins and Families. Dordrecht, the Netherlands, Kluwer Academic, 1992Google Scholar
26. Neale MC, Boker SM, Xie G, Maes HH: Mx: Statistical Modeling, 5th ed. Richmond, Virginia Commonwealth University, Department of Psychiatry, 1999Google Scholar
27. Reich J, Noyes R, Hirschfeld R, Coryell W, O’Gorman TW: State and personality in depressed and panic patients. Am J Psychiatry 1987; 144:181–187Google Scholar
28. Hayward C, Killen JD, Kraemer HC, Taylor CB: Predictors of panic attacks in adolescents. J Am Acad Child Adolesc Psychiatry 2000; 39:207–214Google Scholar
29. Biederman J, Rosenbaum JF, Bolduc-Murphy EA, Faraone SV, Chaloff J, Hirshfeld DR, Kagan J: A 3-year follow-up of children with and without behavioral inhibition. J Am Acad Child Adolesc Psychiatry 1993; 32:814–821Google Scholar
30. Hettema JM, Neale MC, Myers JM, Prescott CA, Kendler KS: A population-based twin study of the relationship between neuroticism and internalizing disorders. Am J Psychiatry 2006; 163:857–864Google Scholar
31. Khan AA, Jacobson KC, Gardner CO, Prescott CA, Kendler KS: Personality and comorbidity of common psychiatric disorders. Br J Psychiatry 2005; 186:190–196Google Scholar
32. Mineka S, Ohman A: Phobias and preparedness: the selective, automatic, and encapsulated nature of fear. Biol Psychiatry 2002; 52:927–937Google Scholar
33. Costa PT, McCrae RR: Revised NEO Personality Inventory (NEO-PI-R) and NEO Five-Factor Inventory (NEO-FFI) Professional Manual. Odessa, Fla, Psychological Assessment Resources, 1992Google Scholar
34. Fullerton J, Cubin M, Tiwari H, Wang C, Bomhra A, Davidson S, Miller S, Fairburn C, Goodwin G, Neale MC, Fiddy S, Mott R, Allison DB, Flint J: Linkage analysis of extremely discordant and concordant sibling pairs identifies quantitative-trait loci that influence variation in the human personality trait neuroticism. Am J Hum Genet 2003; 72:879–890Google Scholar
35. Nash MW, Huezo-Diaz P, Williamson RJ, Sterne A, Purcell S, Hoda F, Cherny SS, Abecasis GR, Prince M, Gray JA, Ball D, Asherson P, Mann A, Goldberg D, McGuffin P, Farmer A, Plomin R, Craig IW, Sham PC: Genome-wide linkage analysis of a composite index of neuroticism and mood-related scales in extreme selected sibships. Hum Mol Genet 2004; 13:2173–2182Google Scholar
36. Stein MB, Schork NJ, Gelernter J: A polymorphism of the beta1-adrenergic receptor is associated with low extraversion. Biol Psychiatry 2004; 56:217–224Google Scholar
37. Kendler KS, Neale MC, Kessler RC, Heath AC, Eaves LJ: A longitudinal twin study of personality and major depression in women. Arch Gen Psychiatry 1993; 50:853–862Google Scholar
38. Maes HH, Neale MC, Kendler KS, Hewitt JK, Silberg JL, Foley DL, Meyer JM, Rutter M, Simonoff E, Pickles A, Eaves LJ: Assortative mating for major psychiatric diagnoses in two population-based samples. Psychol Med 1998; 28:1389–1401Google Scholar
39. Loehlin JC: Genes and Environment in Personality Development. Newbury Park, Calif, Sage Publications, 1992Google Scholar

