
Decreased Prefrontal Cortical Dopamine Transmission in Alcoholism
Abstract
Objective
Basic studies have demonstrated that optimal levels of prefrontal cortical dopamine are critical to various executive functions such as working memory, attention, inhibitory control, and risk/reward decisions, all of which are impaired in addictive disorders such as alcoholism. Based on this and imaging studies of alcoholism that have demonstrated less dopamine in the striatum, the authors hypothesized decreased dopamine transmission in the prefrontal cortex in persons with alcohol dependence.
Method
To test this hypothesis, amphetamine and [11C]FLB 457 positron emission tomography were used to measure cortical dopamine transmission in 21 recently abstinent persons with alcohol dependence and 21 matched healthy comparison subjects. [11C]FLB 457 binding potential, specific compared to nondisplaceable uptake (BPND), was measured in subjects with kinetic analysis using the arterial input function both before and after 0.5 mg kg−1 of d-amphetamine.
Results
Amphetamine-induced displacement of [11C]FLB 457 binding potential (ΔBPND) was significantly smaller in the cortical regions in the alcohol-dependent group compared with the healthy comparison group. Cortical regions that demonstrated lower dopamine transmission in the alcohol-dependent group included the dorsolateral prefrontal cortex, medial prefrontal cortex, orbital frontal cortex, temporal cortex, and medial temporal lobe.
Conclusions
The results of this study, for the first time, unambiguously demonstrate decreased dopamine transmission in the cortex in alcoholism. Further research is necessary to understand the clinical relevance of decreased cortical dopamine as to whether it is related to impaired executive function, relapse, and outcome in alcoholism.
Prefrontal cortical dopamine modulates executive functions such as attention, working memory, and risk/reward decision making (1, 2), all of which are impaired in alcoholism (3–6). Based on this, it is tempting to postulate decreased dopamine transmission in the prefrontal cortex in alcoholism. However, the preclinical literature on this topic is mixed and inconclusive, with studies suggesting increased (7), decreased (8, 9), and no changes (10–12) in prefrontal cortical dopamine transmission in alcoholism. Nevertheless, the ability of prefrontal cortical dopamine to modulate alcohol consumption has been demonstrated in microinjection studies using dopamine D2/3 antagonist and agonist drugs (13, 14). In humans, the displacement of the D2/3 specific positron emission tomography (PET) radiotracer [11C]raclopride following an acute amphetamine (or methylphenidate) challenge has been validated as a noninvasive measure of the change in extracellular dopamine concentration induced by the challenge (15). Using this approach, two research groups have reported decreased striatal dopamine transmission in individuals with alcohol dependence compared with healthy comparison subjects (16, 17). A limitation of these studies was that measurements of dopamine transmission were restricted to the striatum and its subdivisions (i.e., the caudate, putamen, and ventral striatum). Studies were limited to the striatum because [11C]raclopride does not provide sufficient signal-to-noise ratio to quantify D2/3 receptors in extrastriatal areas, such as the cortex, where the concentration of D2/3 receptors is much lower than it is in the striatum. Thus, no previous studies, to our knowledge, have reported on the in vivo status of dopamine in the prefrontal cortex in alcoholism.
We previously validated the high-affinity D2/3 PET radioligand [11C]FLB 457 as a tool to image amphetamine-induced dopamine transmission in the human cortex (18). The results of our validation studies demonstrate 1) low test-retest variability (≤15%) for [11C]FLB 457 binding potential, specific compared to nondisplaceable uptake (BPND), under both baseline and postamphetamine conditions (19, 20); 2) no carryover mass-induced decrease in BPND in the imaging paradigm used to measure dopamine (19); 3) a relatively small fraction of D2/3 receptor specific binding for [11C]FLB 457 in the cerebellar reference region compared with cortical regions of interest (21); and 4) a linear relationship between the amphetamine-induced decreases in [11C]FLB 457 BPND and increases in extracellular dopamine as measured with microdialysis (22). In the present study, we used amphetamine and [11C]FLB 457 PET to contrast cortical dopamine transmission in 21 recently abstinent persons with alcohol dependence and 21 healthy comparison subjects matched on age, gender, race, and nicotine smoking status.
Method
Participants
Seventy-seven individuals with alcohol dependence and 36 healthy comparison subjects were enrolled in the study, and 21 individuals from each group completed the study. The study was approved by the University of Pittsburgh Institutional Review Board and the Radioactive Drug Research Committee. All participants provided written, informed consent. The alcohol-dependent and healthy comparison groups were largely recruited through advertisements displayed at local community centers, on buses, in newspapers, and on websites. Additionally, addiction medicine clinics and hospital emergency departments in the community also referred alcohol-dependent patients. Study criteria for inclusion in the alcohol-dependent group were 1) male or female aged 18–40 years of any ethnic/racial origin; 2) fulfillment of DSM-IV criteria for alcohol dependence as assessed by the Structured Clinical Interview for DSM-IV Axis I Disorders; 3) no current or past DSM-IV axis I disorder other than alcohol abuse or dependence, including abuse of or dependence on other recreational drugs (nicotine dependence was allowed); 4) no current use of cocaine, opiates, cannabis, sedative-hypnotics, amphetamines, 3,4-methylenedioxy-N-methylamphetamine, or phencyclidine (as confirmed by urine drug screen at the study screening); 5) not currently taking any prescription or over-the-counter medications; 6) no current or past chronic medical or neurological illnesses (including glaucoma, seizure disorders, a focal finding on MRI such as stroke or tumor, chronic obstructive pulmonary disease or respiratory disease, renal problems, and liver problems) as assessed by a complete physical examination and laboratory examination; 7) no resting systolic blood pressure >140 mm Hg and no diastolic blood pressure >90 mm Hg; 8) no more than one risk factor for coronary artery disease (e.g., smoking, obesity, cholesterol >240 mg dl−1, sedentary lifestyle, etc.); 9) no first-degree relative with a psychotic or mood disorder; 10) not currently pregnant; 11) no history of radioactivity exposure from nuclear medicine studies or occupation; and 12) no metallic objects in the body that are contraindicated for MRI.
Participants in the alcohol-dependent group completed a minimum of 14 days of outpatient abstinence monitored with witnessed urine toxicology. They were monitored for alcohol and recreational drug use with urine alcohol metabolites (ethyl glucuronide and ethyl sulfate) and urine drug screens three times per week for 2 consecutive weeks. Since alcohol metabolites and common drugs of abuse can be detected for 2–3 days after use, individuals in this group were informed that they should not use alcohol or street drugs for the 14 days before the PET study. In order to promote abstinence from alcohol during this 2-week period, they were paid $75.00 for each urine sample that was negative for ethyl glucuronide and ethyl sulfate. They were scheduled for the PET scans after successful completion of the abstinence monitoring protocol. Individuals who tested positive for ethyl glucuronide and ethyl sulfate were offered up to three attempts to restart the abstinence monitoring protocol. This abstinence monitoring protocol ensured that all persons in the alcohol-dependent group were abstinent for a minimum of 2 weeks before the PET scan. This group was also monitored for alcohol withdrawal signs and symptoms three times during the first week of abstinence using the Clinical Institute Withdrawal Assessment for Alcohol Scale (23). Those who were at risk of severe withdrawal (i.e., scored >19 on the Clinical Institute Withdrawal Assessment for Alcohol Scale and had a history of alcohol withdrawal seizures or delirium tremens) were excluded from the research protocol. The severity of alcohol dependence was assessed with the Michigan Alcoholism Screening Test (24) and the Alcohol Dependence Scale (25).
Healthy comparison subjects, matched for age, gender, ethnicity, and smoking status, had no past or present neurological or psychiatric illnesses, including substance abuse (as confirmed by urine drug screen, at both the study screening and the day of the PET scan). Both the healthy comparison and alcohol-dependent groups underwent the PET scans as outpatients. Following the completion of the PET scans, those in the alcohol-dependent group were scheduled for a follow-up visit, during which they were debriefed of the risks of alcohol abuse and provided with a referral for outpatient treatment.
Image Acquisition and Analysis
Following a structural MRI, participants underwent a baseline and a postamphetamine [11C]FLB 457 PET scan in the same experimental session using procedures previously described (18).
Briefly, [11C]FLB 457 was synthesized using the methodology reported by Halldin et al. (26). PET imaging sessions were conducted with the ECAT EXACT HR Plus camera (CTI-Siemens, Munich, Germany). Following a transmission scan, participants received an intravenous bolus injection of [11C]FLB 457, and emission data were collected for 90 minutes. Arterial blood samples were collected to measure the plasma free fraction (fP) for [11C]FLB 457 and to derive a metabolite-corrected arterial input function for modeling using methods previously described (18). The maximum injected mass for [11C]FLB 457 was restricted to 0.6 μg (27). The postamphetamine [11C]FLB 457 scan was performed 3 hours after the administration of 0.5 mg kg−1 of oral d-amphetamine. During this scan, amphetamine blood levels were measured in three arterial samples drawn at 0 minutes, 45 minutes, and 90 minutes and analyzed using methods described by Reimer et al. (28).
PET data were reconstructed and processed with the image analysis software MEDx (Sensor Systems, Sterling, Va.) and SPM2 (www.fil.ion.ucl.ac.uk/spm) as previously described (18). Frame-to-frame motion correction for head movement and MR-PET image alignment were performed using a mutual information algorithm implemented in SPM2. MRI segmentation was performed using the automated segmentation tool in Functional MRI of the Brain Software Library (29). Cortical (medial temporal lobe, dorsolateral prefrontal cortex, orbital frontal cortex, medial prefrontal cortex, anterior cingulate cortex, temporal cortex, parietal cortex, and occipital cortex) and subcortical (midbrain and cerebellum) regions of interest were defined on the MRI using a segmentation-based and direct identification method previously described (19, 30, 31). Regional volumes and time activity curves were then generated in MEDx as outlined by Abi-Dargham et al. (30, 31). Primary analysis included the eight cortical regions that had been validated in our previous [11C]FLB 457 human studies (18–21). Secondary analysis included the midbrain as a region of interest to test whether there is convergence between the midbrain dopamine cells and terminal fields. Derivation of [11C]FLB 457 volumes of distribution expressed relative to total plasma ligand concentration (VT) in the regions of interest (VT ROI) and cerebellum (VT CER) was performed using a two-tissue compartment kinetic analysis with the arterial input function we previously described (18).
PET outcome variables are defined in accordance to the consensus nomenclature for in vivo imaging of reversibly binding radioligands (32). The D2/3 receptor availability at the baseline and postamphetamine conditions was estimated using BPND, which was derived as follows:

In this equation, fND (which equals fP/VT CER) is the free fraction of [11C]FLB 457 in the nondisplaceable compartment, Bavail is the density of D2/3 receptors (nmol L−1) available to bind to [11C]FLB 457 in vivo, and KD is the in vivo equilibrium dissociation constant of [11C]FLB 457 (nmol L−1).
The amphetamine-induced change in BPND (ΔBPND) was calculated as the difference between BPND measured in the postamphetamine condition (BPND AMPH) and BPND measured in the baseline condition (BPND BASE) and expressed as a percentage of BPND BASE as follows:

Finally, because our validation work with [11C]FLB 457 demonstrated D2/3 specific binding in the cerebellum, there was concern that any amphetamine-induced change in VT CER could bias the dopamine-release outcome measure, ΔBPND (BPND is dependent on VT CER; see Equation 1). Therefore, to arrive at a dopamine-release measurement in the cortex (i.e., D2/3 receptor occupancy following amphetamine administration) that is independent of VT CER, we analyzed the baseline and postamphetamine VT values from the eight cortical regions of interest with Lassen plots as described by Cunningham et al. (33). Briefly, the equation for the line [y=mx+b], where y=[VT BASELINE–VT AMPHETAMINE] and x=VT BASELINE, produced a linear relationship with the slope of line equal to the receptor occupancy (m). This approach assumed that there is uniform receptor occupancy across the cortical regions.
Statistical Analysis
Comparisons between scan conditions (baseline and postamphetamine) were performed with paired t tests. Comparisons between groups (alcohol-dependent and healthy comparison subjects) were performed with unpaired t tests (regions-of-interest level) and repeated-measures of analysis of variance (ANOVA) (amphetamine blood levels). Furthermore, to test for a global effect of diagnosis (alcohol-dependent and healthy comparison) on baseline cortical BPND and ΔBPND, a linear mixed-model analysis was performed with cortical regions of interest as a repeated measure and diagnostic group as the fixed factor (IBM SPSS Statistics, IBM, Armonk, N.Y.). Relationships between PET data and clinical characteristics (years of drinking, amount of drinks consumed per day, and Alcohol Dependence Scale and Michigan Alcohol Screening Test scores) were assessed using the Pearson product-moment correlation coefficient. A two-tailed probability (p) value <0.05 was selected as the significance level for all analyses. A false discovery rate correction with an alpha set at 0.05 was applied to correct for multiple comparisons in the regions of interest (34).
Results
Twenty-one individuals with alcohol dependence were matched with 21 healthy comparison subjects on age, gender, ethnicity, and smoking status (including the number of cigarettes smoked per day rounded to the nearest one-half pack, i.e., 10 cigarettes). The demographic variables for both groups and the measures of alcohol use in the alcohol-dependent group are summarized in Table 1.
| Characteristic | Healthy Comparison Group (N=21) | Alcohol-Dependent Group (N=21) | ||
|---|---|---|---|---|
| N | % | N | % | |
| Gender | ||||
| Male | 16 | 76.2 | 16 | 76.2 |
| Female | 5 | 23.8 | 5 | 23.8 |
| Ethnicity | ||||
| African American | 2 | 9.5 | 4 | 19.0 |
| Asian | 2 | 9.5 | 0 | 0.0 |
| Caucasian | 17 | 81.0 | 17 | 81.0 |
| Smoking status | ||||
| Yes | 12 | 57.1 | 12 | 57.1 |
| No | 9 | 42.9 | 9 | 42.9 |
| Positive family history for alcoholism | 0 | 0.0 | 18 | 85.7 |
| Mean | SD | Mean | SD | |
| Age (years) | 28 | 4 | 28 | 5 |
| Weight (kg) | 74 | 12 | 77 | 14 |
| Duration of abuse (years) | 11 | 6 | ||
| Alcohol frequency (days per week) | 5.7 | 1.5 | ||
| Alcohol amount (standard number of drinks per day) | 13 | 5 | ||
| Michigan Alcohol Screening Test score (range: 0–22) | 13 | 4 | ||
| Alcohol Dependence Scale score (range: 0–47) | 21 | 6 | ||
| Abstinence before scans (days) | 33 | 18 | ||
Scan Parameters
The [11C]FLB 457 scan parameters for the healthy comparison and alcohol-dependent groups under baseline and postamphetamine conditions are presented in Table 2. No significant differences between the baseline and postamphetamine conditions were observed in any of these scan parameters in either group. Notably, there was no significant change in VT CER following amphetamine administration in both groups (ΔVT CER: healthy comparison=−2.5% [SD=15.1]; alcohol-dependent=−0.9% [SD=10.8]; t=0.41, df=40, p=0.68). This justified the use of ΔBPND to contrast differences in amphetamine-induced dopamine transmission between the two groups (see Equation 1, also see reference 35).
| Group and Condition | Injected Dose (mCi) | Specific Activity (Ci mmol–1) | Injected Mass (μg) | Free Fraction in Plasma (%) | Nondisplaceable Volume of Distribution (mL cm–3) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Healthy comparison | ||||||||||
| Baseline | 7.1 | 1.7 | 8,891 | 5,689 | 0.39 | 0.17 | 34.7 | 5.9 | 4.27 | 1.23 |
| Postamphetamine | 7.7 | 1.4 | 8,741 | 4,307 | 0.39 | 0.13 | 34.3 | 5.1 | 4.04 | 0.86 |
| Alcohol-dependent | ||||||||||
| Baseline | 7.6 | 1.4 | 11,369 | 8,871 | 0.36 | 0.17 | 38.9 | 7.6 | 4.43 | 1.01 |
| Postamphetamine | 6.8 | 1.9 | 6,995 | 4,736 | 0.46 | 0.16 | 39.3 | 7.3 | 4.37 | 0.97 |
The amphetamine blood levels measured at 0, 45, and 90 minutes during the postamphetamine [11C]FLB 457 scan in the healthy comparison group (80 ng mL−1 [SD=10], 73 ng mL−1 [SD=9], and 70 ng mL−1 [SD=10]) were not significantly different from the amphetamine levels measured in the alcohol-dependent group (75 ng mL−1 [SD=8], 69 ng mL−1 [SD=7], and 66 ng mL−1 [SD=7]; repeated measures of ANOVA, effect of diagnosis: F=3.08, df=1, 40, p=0.09; effect of time: F=57.20, df=2, 39, p<0.001; diagnosis-by-time interaction: F=0.07, df=2, 39, p=0.93).
Regional Volumes
No between-group differences were found in volumes in the cortical regions, midbrain, and cerebellum determined from the MRI scans, suggesting a lack of measurable volumetric changes in alcohol dependence.
Cortex
D2/3 receptor availability (BPND) under baseline conditions.
As shown in Figure 1, no differences in baseline [11C]FLB 457 BPND were observed in the alcohol-dependent group compared with the healthy comparison group (linear mixed model: effect of diagnosis: F=0.89, df=1, 40, p=0.35; effect of region: F=332.65, df=7, 280, p<0.001; region-by-diagnosis interaction: F=1.71, df=7, 280, p=0.11). Additionally, unpaired t tests conducted at the level of the individual regions of interest did not show any significant differences between the two groups.
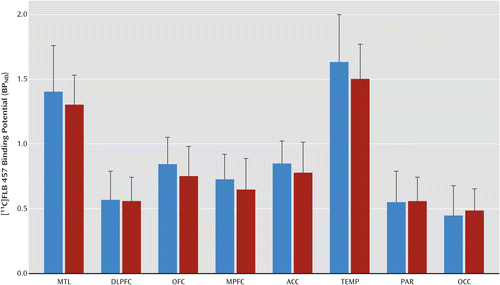
a The bar graph shows the lack of difference in D2/3 receptor availability in cortical regions of interest in the alcohol-dependent group (red bars) compared with the healthy comparison group (blue bars). MTL=medial temporal lobe; DLPFC=dorsolateral prefrontal cortex; OFC=orbital frontal cortex; MPFC=medial prefrontal cortex; ACC=anterior cingulate cortex; TEMP=temporal cortex; PAR=parietal cortex; and OCC=occipital cortex.
Amphetamine-induced reduction in D2/3 receptor availability (ΔBPND).
Amphetamine led to a significant reduction in [11C]FLB 457 BPND in healthy comparison subjects (Figure 2A) but not in those with alcohol dependence (Figure 2B). The amphetamine-induced Δ[11C]FLB 457 BPND was significantly lower in the alcohol-dependent group compared with the healthy comparison group (linear mixed model: effect of diagnosis: F=11.03, df=1, 40, p=0.002; effect of region: F=1.99, df=7, 280, p<0.056; region-by-diagnosis interaction: F=0.65, df=7, 280, p=0.71). The inclusion of mean amphetamine blood levels as a covariate in the model did not change the significance of the results (linear mixed model: effect of diagnosis: F=9.44, df=1, 39, p=0.004). Unpaired t tests conducted at the level of the individual regions of interest were significant in all of the cortical regions except the anterior cingulate cortex (Table 3). All significant comparisons, except in the parietal and occipital cortices, survived the false discovery rate correction. Furthermore, consistent with the ΔBPND results, the amphetamine-induced dopamine release occupancy of D2/3 receptors (derived using Lassen plots independent of VT CER) was significantly lower in the alcohol-dependent group compared with the healthy comparison group (healthy comparison=16.0% [SD=15.6]; alcohol-dependent=−1.2% [SD=19.5]; t=−3.16, df=40, p=0.003).
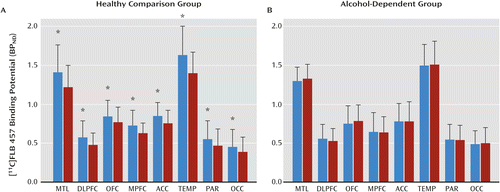
a The bar graphs show [11C]FLB 457 BPND under baseline (blue bars) and postamphetamine (red bars) conditions in the healthy comparison group (panel A) and the alcohol-dependent group (panel B). Amphetamine led to a significant decrease in [11C]FLB 457 BPND in healthy comparison subjects but not in the alcohol-dependent group ([*] represents p<0.05, following the false discovery rate correction for multiple comparisons). MTL=medial temporal lobe; DLPFC=dorsolateral prefrontal cortex; OFC=orbital frontal cortex; MPFC=medial prefrontal cortex; ACC=anterior cingulate cortex; TEMP=temporal cortex; PAR=parietal cortex; and OCC=occipital cortex.
| Region | Healthy Comparison Group (N=21) | Alcohol-Dependent Group (N=21) | Analysisa | ||||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | t | df | p | |
| Medial temporal lobe | –11.1 | 16.2 | 3.1 | 15.9 | 2.88 | 39.98 | 0.006b |
| Dorsolateral prefrontal cortex | –13.7 | 14.7 | –3.9 | 12.7 | 2.30 | 39.12 | 0.03b |
| Orbital frontal cortex | –8.6 | 12.4 | 8.5 | 18.6 | 3.50 | 34.87 | 0.001b |
| Medial prefrontal cortex | –11.6 | 15.1 | 5.4 | 27.3 | 2.48 | 31.62 | 0.02b |
| Anterior cingulate cortex | –9.0 | 17.7 | 1.1 | 19.6 | 1.75 | 39.61 | 0.09 |
| Temporal cortex | –12.2 | 13.4 | 0.4 | 10.4 | 3.39 | 37.80 | 0.002b |
| Parietal cortex | –11.7 | 18.6 | –1.6 | 12.2 | 2.09 | 34.65 | 0.04 |
| Occipital cortex | –10.6 | 19.2 | 1.3 | 17.2 | 2.11 | 39.52 | 0.04 |
Midbrain
D2/3 receptor availability (BPND) under baseline conditions.
No significant differences were observed in the midbrain [11C]FLB 457 BPND in the alcohol-dependent group compared with the healthy comparison group (BPND: healthy comparison=2.50 [SD=0.62]; alcohol-dependent=2.23 [SD=0.38]; t=−1.67, df=40, p=0.10).
Amphetamine-induced reduction in D2/3 receptor availability (ΔBPND)
Amphetamine led to a significant reduction in [11C]FLB 457 BPND in the midbrain in the healthy comparison group (baseline=2.50 [SD=0.62]; postamphetamine=2.09 [SD=0.37]; t=3.16, df=20, p=0.005) but not in the alcohol-dependent group (baseline=2.23 [SD=0.38]; postamphetamine=2.35 [SD=0.63]; t=−1.49, df=20, p=0.15). The amphetamine-induced Δ[11C]FLB 457 BPND in the midbrain was significantly lower in the alcohol-dependent group compared with the healthy comparison group (ΔBPND: healthy comparison=−13.2% [SD=15.2]; alcohol-dependent=5.0% [SD=14.6]; t=3.91, df=40, p=0.0003). This comparison survived the false discovery rate correction.
Clinical Correlations
Correlation analyses revealed no significant associations between ΔBPND in the regions of interest (the cortex and midbrain) and mean amphetamine blood levels in the healthy comparison and alcohol-dependent groups. Additionally, there was no significant association between ΔBPND and any of the clinical measures (alcohol frequency, amount of consumption, duration of abuse, and Michigan Alcohol Screening Test and Alcohol Dependence Scale scores) in the alcohol-dependent group.
Discussion
In this PET study, we found less displacement of [11C]FLB 457 BPND in the cortex and midbrain after amphetamine administration in recently abstinent alcohol-dependent subjects compared with healthy comparison subjects. In a previous study using PET and microdialysis, it was shown that 1% displacement of [11C]FLB 457 BPND in the cortex corresponds to a 57% increase in extracellular dopamine concentration (22). Extending this relationship to the present data set (mean ΔBPND: healthy comparison group: −9% to −14%; alcohol-dependent group: –4% to +9% [Table 3]) suggests that cortical dopamine in the healthy comparison and alcohol-dependent subjects increases by approximately 513%−798% and 0%−228%, respectively, following the same dose of amphetamine. This result, for the first time to our knowledge, unequivocally demonstrates that there is decreased dopamine transmission in the cortex in alcoholism. Also for the first time, these data show convergence between the midbrain dopamine cells and terminal fields with respect to decreased dopamine transmission in alcoholism. Such a blunting in mesocortical dopamine transmission in persons with alcohol dependence is consistent with previously reported findings in the nigrostriatal system that includes the limbic-related nucleus accumbens (16). Decreased dopamine transmission in the mesolimbic regions, such as the ventral striatum and medial temporal lobe, likely contributes to anhedonia, amotivation, and decreased reward sensitivity in alcohol dependence. This has led to the conceptualization of alcohol dependence as a reward-deficit disorder with a higher reward threshold for both natural and drug/alcohol reinforcers (36, 37). The fact that there is also less dopamine in the prefrontal cortex, which governs executive functions, is important because it could impair the addicted person’s ability to learn and utilize informational/behavioral strategies critical to relapse prevention. This is supported by literature that links prefrontal cortical dopamine with executive functions, such as attention, working memory, behavioral flexibility, and risk/reward decision making, all of which are impaired in addictive disorders such as alcoholism (3, 38). In a study using a rodent version of the Iowa Gambling Task (a task that measures risk-preference decision making), Floresco and Magyar (38) demonstrated that blocking dopamine transmission in the prefrontal cortex leads to a response decision that fails to integrate the consequences of conditioned punishment. Based on this study, it is tempting to speculate that the failure to incorporate past negative consequences in a decision to drink alcohol during abstinence is related to decreased prefrontal cortical dopamine in alcoholism. If this hypothesis were confirmed, it would support a role for medications that increase prefrontal cortical dopamine to prevent relapse in alcoholism.
Alcohol-induced potentiation of GABA, the major inhibitory transmitter in the brain, inhibits GABA-ergic interneurons in the ventral tegmental area and substantia nigra and leads to increased phasic (or synaptic) dopamine transmission (39, 40). However, chronic and repeated use of alcohol leads to decreased phasic dopamine through adaptations in the tonic (or extracelluar) dopamine and glutamatergic systems in the cortico-limbic pathways (for a detailed review, see reference 39). If decreased dopamine transmission is the result of an adaptation in the cortico-limbic circuits, it might be possible to reverse this deficit in persons with alcohol dependence with prolonged abstinence. On the other hand, alcohol-dependent persons also demonstrate signs of inflammation (i.e., greater activated microglia and proinflammatory cytokines) and a reduction of dopamine neuronal markers in the brain (41, 42). Therefore, the possibility of a toxic irreversible loss of dopamine neurons in alcoholism cannot be ruled out. This may explain the persistent and enduring cognitive impairments that have been reported in abstinent persons with alcohol dependence (43). Also unclear is whether decreased dopamine transmission in alcoholism represents a premorbid trait or alcohol-induced state. Future dopamine imaging studies of individuals recovering from alcoholism with prolonged periods of abstinence and nonhuman primates that can be imaged both pre- and postalcohol exposure are necessary to evaluate these issues.
Another interesting observation in the present study is the lack of differences in baseline D2/3 receptor binding potential in both the cortex and midbrain in the alcohol-dependent group compared with the healthy comparison group (Figure 1). This is in contrast to previous [11C]raclopride imaging studies that have reported a decrease in D2/3 receptor BPND of approximately 10%−20% in the striatal subdivisions in persons with alcohol dependence (16). The exact reason and physiological relevance for decreased D2/3 receptor BPND in the striatum, but not in the extrastriatal regions, in alcoholism is not clear. Reasons that may have contributed to the inability to detect group differences in our study include diminished power because of greater between-subject variability in cortical D2/3 receptor binding potential and/or a more pronounced reduction in baseline dopamine levels (i.e., before amphetamine stimulation) in the cortex compared with in the striatum in the alcohol-dependent group. PET studies in alcohol-dependent individuals using alpha-methyl-para-tyrosine that can deplete baseline dopamine in the striatal and extrastriatal regions are necessary to further understand this issue.
The strengths of this study are inclusion of relatively young individuals (≤40 years old) with mild to moderate alcohol dependence; exclusion of individuals with comorbid medical illness, psychiatric illness, or drug abuse; monitoring of abstinence before imaging; use of a validated imaging paradigm to measure cortical dopamine transmission; use of compartmental modeling with an arterial input function to derive PET outcome measures; ruling out changes in [11C]FLB 457 nonspecific binding (nondisplaceable volume of distribution) as a significant contributor to ΔBPND in both alcohol-dependent and healthy comparison subjects; and measurement of amphetamine blood levels. However, this study does have some limitations, including the exclusion of older individuals with severe alcohol dependence and no identification of a relationship between ΔBPND and alcohol measures such as the frequency, amount, severity, and duration of abuse. One possible reason for the failure to demonstrate a relationship between ΔBPND and alcohol measures is the limited range of values observed in the alcohol abuse measurements (see Table 1). This is likely an unintended consequence of excluding individuals with more severe alcoholism and comorbid disorders. In conclusion, we found decreased dopamine transmission in abstinent persons with alcohol dependence in several of the cortical regions that have been implicated in addiction, including the prefrontal cortex and medial temporal lobe. The results of this study suggest that dopamine dysfunction in alcohol dependence is more widespread than previously conceptualized and not restricted to the striatum. Further studies are necessary to understand the mechanisms that contribute to blunted dopamine transmission and its clinical relevance in alcoholism.
1 : Cognitive deficit caused by regional depletion of dopamine in prefrontal cortex of rhesus monkey. Science 1979; 205:929–932Crossref, Medline, Google Scholar
2 : D1 dopamine receptors in prefrontal cortex: involvement in working memory. Science 1991; 251:947–950Crossref, Medline, Google Scholar
3 : Are executive function and impulsivity antipodes? a conceptual reconstruction with special reference to addiction. Psychopharmacology (Berl) 2012; 221:361–387Crossref, Medline, Google Scholar
4 : The effect of intellectual deterioration on retention deficits in amnesic alcoholics. J Abnorm Psychol 1988; 97:448–454Crossref, Medline, Google Scholar
5 : Cognitive impairment in chronic alcoholics: some cause for optimism. Am Psychol 1983; 38:1045–1054Crossref, Medline, Google Scholar
6 : Impairments of brain and behavior: the neurological effects of alcohol. Alcohol Health Res World 1997; 21:65–75Medline, Google Scholar
7 : Sardinian alcohol-preferring rats show low 5-HT extraneuronal levels in the mPFC and no habituation in monoaminergic response to repeated ethanol consumption in the NAcS. Brain Res 2004; 1006:18–27Crossref, Medline, Google Scholar
8 : Extracellular dopamine levels are lower in the medial prefrontal cortex of alcohol-preferring rats compared to Wistar rats. Alcohol 2006; 38:5–12Crossref, Medline, Google Scholar
9 : Phenotypic and genotypic characterization of the Indiana University rat lines selectively bred for high and low alcohol preference. Behav Genet 2002; 32:363–388Crossref, Medline, Google Scholar
10 : Non-psychostimulant drugs of abuse and anxiogenic drugs activate with differential selectivity dopamine transmission in the nucleus accumbens and in the medial prefrontal cortex of the rat. Psychopharmacology (Berl) 1996; 124:293–299Crossref, Medline, Google Scholar
11 : Modulation of the stress response by ethanol in the rat frontal cortex. Pharmacol Biochem Behav 1993; 45:327–334Crossref, Medline, Google Scholar
12 : Effects of acute ethanol administration on monoamine and metabolite content in forebrain regions of ethanol-tolerant and nontolerant alcohol-preferring (P) rats. Pharmacol Biochem Behav 1988; 29:169–174Crossref, Medline, Google Scholar
13 : Dopamine receptors in the medial prefrontal cortex influence ethanol and sucrose-reinforced responding. Alcohol Clin Exp Res 1996; 20:1631–1638Crossref, Medline, Google Scholar
14 : Dopaminergic involvement in medial prefrontal cortex and core of the nucleus accumbens in the regulation of ethanol self-administration: a dual-site microinjection study in the rat. Physiol Behav 2003; 79:581–590Crossref, Medline, Google Scholar
15 : Schizophrenia is associated with elevated amphetamine-induced synaptic dopamine concentrations: evidence from a novel positron emission tomography method. Proc Natl Acad Sci USA 1997; 94:2569–2574Crossref, Medline, Google Scholar
16 : Profound decreases in dopamine release in striatum in detoxified alcoholics: possible orbitofrontal involvement. J Neurosci 2007; 27:12700–12706Crossref, Medline, Google Scholar
17 : Alcohol dependence is associated with blunted dopamine transmission in the ventral striatum. Biol Psychiatry 2005; 58:779–786Crossref, Medline, Google Scholar
18 : Positron emission tomography imaging of amphetamine-induced dopamine release in the human cortex: a comparative evaluation of the high affinity dopamine D2/3 radiotracers [11C]FLB 457 and [11C]fallypride. Synapse 2009; 63:447–461Crossref, Medline, Google Scholar
19 : Positron emission tomography imaging of dopamine D2/3 receptors in the human cortex with [¹¹C]FLB 457: reproducibility studies. Synapse 2011; 65:35–40Crossref, Medline, Google Scholar
20 : Reproducibility of post-amphetamine [11C]FLB 457 binding to cortical D2/3 receptors. PLoS ONE 2013; 8:e76905Crossref, Medline, Google Scholar
21 : Evaluation of dopamine D2/3 specific binding in the cerebellum for the positron emission tomography radiotracer [¹¹C]FLB 457: implications for measuring cortical dopamine release. Synapse 2011; 65:991–997Crossref, Medline, Google Scholar
22 : Imaging dopamine transmission in the frontal cortex: a simultaneous microdialysis and [11C]FLB 457 PET study. Mol Psychiatry 2014; 19:302–310Crossref, Medline, Google Scholar
23 : Assessment of alcohol withdrawal: the Revised Clinical Institute Withdrawal Assessment for Alcohol Scale (CIWA-Ar). Br J Addict 1989; 84:1353–1357Crossref, Medline, Google Scholar
24 : The Michigan Alcoholism Screening Test: the quest for a new diagnostic instrument. Am J Psychiatry 1971; 127:1653–1658Link, Google Scholar
25 : Alcohol dependence syndrome: measurement and validation. J Abnorm Psychol 1982; 91:199–209Crossref, Medline, Google Scholar
26 : Carbon-11-FLB 457: a radioligand for extrastriatal D2 dopamine receptors. J Nucl Med 1995; 36:1275–1281Medline, Google Scholar
27 : Reproducibility of [11C]FLB 457 binding in extrastriatal regions. Nucl Med Commun 2001; 22:1215–1221Crossref, Medline, Google Scholar
28 : Determination of amphetamine, methamphetamine and desmethyldeprenyl in human plasma by gas chromatography/negative ion chemical ionization mass spectrometry. Biol Mass Spectrom 1993; 22:235–242Crossref, Medline, Google Scholar
29 : Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004; 23(suppl 1):S208–S219Crossref, Medline, Google Scholar
30 : Measurement of striatal and extrastriatal dopamine D1 receptor binding potential with [11C]NNC 112 in humans: validation and reproducibility. J Cereb Blood Flow Metab 2000; 20:225–243Crossref, Medline, Google Scholar
31 : Prefrontal dopamine D1 receptors and working memory in schizophrenia. J Neurosci 2002; 22:3708–3719Crossref, Medline, Google Scholar
32 : Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab 2007; 27:1533–1539Crossref, Medline, Google Scholar
33 : Measuring drug occupancy in the absence of a reference region: the Lassen plot re-visited. J Cereb Blood Flow Metab 2010; 30:46–50Crossref, Medline, Google Scholar
34 : Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc 1995; 57:289–300Google Scholar
35 : A comparative evaluation of the dopamine D2/3 agonist radiotracer [11C]NPA and antagonist [11C]raclopride to measure amphetamine-induced dopamine release in the human striatum. J Pharmacol Exp Ther 2010; 63:574–584Google Scholar
36 : Neurocircuitry of addiction. Neuropsychopharmacology 2010; 35:217–238Crossref, Medline, Google Scholar
37 : Theoretical frameworks and mechanistic aspects of alcohol addiction: alcohol addiction as a reward deficit disorder. Curr Top Behav Neurosci 2013; 13:3–30Crossref, Medline, Google Scholar
38 : Mesocortical dopamine modulation of executive functions: beyond working memory. Psychopharmacology (Berl) 2006; 188:567–585Crossref, Medline, Google Scholar
39 : The tonic/phasic model of dopamine system regulation and its implications for understanding alcohol and psychostimulant craving. Addiction 2000; 95(suppl 2):S119–S128Crossref, Medline, Google Scholar
40 : Sex differences in striatal dopamine release in young adults after oral alcohol challenge: a positron emission tomography imaging study with [¹¹C]raclopride. Biol Psychiatry 2010; 68:689–696Crossref, Medline, Google Scholar
41 : Increased MCP-1 and microglia in various regions of the human alcoholic brain. Exp Neurol 2008; 210:349–358Crossref, Medline, Google Scholar
42 : Decreased striatal monoaminergic terminals in severe chronic alcoholism demonstrated with (+)[11C]dihydrotetrabenazine and positron emission tomography. Ann Neurol 1998; 44:326–333Crossref, Medline, Google Scholar
43 : Cognitive functions in abstinent alcohol-dependent patients. Alcohol 2012; 46:665–671Crossref, Medline, Google Scholar

