
Evidence That Schizophrenia Risk Variation in the ZNF804A Gene Exerts Its Effects During Fetal Brain Development
Abstract
Objective
The single-nucleotide polymorphism rs1344706, located within an intron of the ZNF804A gene, exhibits genome-wide significant association with schizophrenia. Although genotype at rs1344706 is associated with altered functional brain connectivity, the molecular mechanisms mediating its association with schizophrenia have not been clearly defined. Given its location in noncoding sequence, the authors tested association between rs1344706 and ZNF804A expression in adult and fetal human brain.
Method
Highly quantitative measures of relative allelic expression were used to assess the effect of rs1344706 genotype on the mRNA expression of ZNF804A in the dorsolateral prefrontal cortex, hippocampus, and substantia nigra of the adult human brain and in human brain tissue from the first and second trimester of gestation.
Results
Genotype at rs1344706 had no significant effect on the regulation of ZNF804A in any of the three adult brain regions examined. In contrast, rs1344706 genotype had a significant effect on ZNF804A allelic expression in second-trimester fetal brain, with the schizophrenia risk (T) allele associated with reduced ZNF804A expression.
Conclusions
The T allele of rs1344706 is associated with a relative decrease in ZNF804A expression during the second trimester of fetal brain development. These data provide evidence for a schizophrenia risk mechanism that is operational in (and possibly specific to) the fetal brain.
Large genome-wide association studies (GWASs) have identified several novel susceptibility loci for psychiatric disorders (1, 2). The first genome-wide significant association with the broad phenotype of psychosis (encompassing both schizophrenia and bipolar disorder) was at the single-nucleotide polymorphism (SNP) rs1344706, which is located within an intron of the zinc finger protein 804A (ZNF804A) gene (3). Although association with rs1344706 was not reported to be one of the most significant findings in a recent mega-analysis of schizophrenia GWAS data (4), the association of the T allele of this SNP with schizophrenia is supported by several independent replication studies (5–7), and in a meta-analysis of more than 55,000 subjects (18,945 schizophrenia/schizoaffective disorder case subjects and 38,675 comparison subjects) this association greatly exceeded accepted levels of genome-wide significance (p=2.5×10−11) (8). There is also evidence for additional schizophrenia risk variants in the ZNF804A gene (9, 10). However, extensive resequencing of the ZNF804A locus in samples from the United Kingdom did not uncover a variant that exhibits stronger association with schizophrenia than rs1344706 (8).
Functional MRI (fMRI) studies have demonstrated that genotype at rs1344706 is associated with altered functional connectivity between the prefrontal cortex and hippocampus (11, 12). However, the molecular mechanisms that mediate association between this SNP and psychiatric illness have not been clearly defined. We previously demonstrated that the schizophrenia-associated T allele of rs1344706 results in reduced binding of an unidentified nuclear protein in neural cells (13), suggesting direct effects of this SNP on ZNF804A expression or splicing. However, while rs1344706 has been reported to exhibit association with ZNF804A expression in adult tissues (5, 8), this association appears to be through additional regulatory variants at the ZNF804A locus rather than through direct effects of rs1344706 (8).
Genetic variants that affect a gene’s expression by altering its regulatory sequences are said to be acting in cis. Variable cis effects on gene expression can be readily detected by comparing the expression of one chromosomal copy of the gene with the other in individual subjects (14, 15). This is because a cis regulatory variant will only affect expression of the gene copy that is transcribed from the parental chromosome on which it resides. In contrast, trans effects on gene expression arise from remote sources (e.g., other gene loci) and act on both chromosomal copies of the regulated gene.
The allelic expression approach typically makes use of a SNP in exonic (i.e., expressed) sequence to discriminate the RNA transcribed from each chromosomal copy of a gene in heterozygous individuals. This allows the net effects of regulatory variants operating on each allele to be relatively quantified while holding constant trans-acting influences on gene expression arising from other gene loci or environmental factors. In the absence of genetic or epigenetic variants affecting the expression of a gene in cis, the two chromosomal copies of that gene will be expressed equally. Conversely, departure from the expected 1:1 ratio of allelic expression in any given sample indicates heterozygous cis regulatory influences operating on that gene (14, 15). Because assays of allelic expression expose only cis effects on gene expression, they offer a powerful means of testing association between particular DNA variants and variable cis regulation of the predicted target gene. Such effects are indicated when allelic expression ratios in genomic heterozygotes for the variant differ from those in genomic homozygotes for that variant, since it is only in heterozygotes that any cis effects of the variant will be unequal (16, 17).
It has previously been demonstrated that allelic expression can differ between tissue, cell types, and developmental stages (18–21). This may be explained by cellular differences in trans-acting factors (e.g., transcription factors) that bind to sequences containing cis regulatory variants and thereby elicit their effects (22). Such observations suggest that allelic expression measures can be used to test not only whether identified genetic risk variants affect the cis regulation of a gene, but also where and when they are active. In this study, we used measures of allelic expression to test potential effects of rs1344706 on the cis regulation of ZNF804A in three regions of the adult brain and at two stages of fetal brain development.
Method
Brain Samples
Postmortem adult human brain tissue was obtained from the Medical Research Council London Neurodegenerative Diseases Brain Bank. Ethical approval for our study was obtained from the Joint South London and Maudsley and the Institute of Psychiatry National Health Service Research Ethics Committee. Adult brain samples were selected from an initial sample of 96 nonrelated subjects on the basis of heterozygosity for the ZNF804A-expressed SNP rs12476147 (used to measure ZNF804A allelic expression) and availability of tissue from at least one of three chosen brain regions (the dorsolateral prefrontal cortex, hippocampus, and substantia nigra). All subjects had no psychiatric or neurological diagnosis at the time of death. Assayed dorsolateral prefrontal cortex (Brodmann’s area 9) samples (heterozygous for rs12476147 and therefore informative for the allelic expression assay) were derived from 32 subjects, with an average age at death of 72 years (range: 20–102 years) and 68 years (range: 40–89 years) for rs1344706 heterozygotes and homozygotes, respectively. Assayed hippocampal samples (heterozygous for rs12476147) were derived from 21 subjects, with an average age at death of 70 years (range: 20–102 years) and 66 years (range: 40–89 years) for rs1344706 heterozygotes and homozygotes, respectively. Assayed substantia nigra samples (heterozygous for rs12476147) were derived from 18 subjects, with an average age at death of 69 years (range: 20–96 years) and 68 years (range: 43–89 years) for rs1344706 heterozygotes and homozygotes, respectively. Tissue was derived from all three brain regions from 14 subjects. There was no significant difference in age between heterozygotes and homozygotes in any brain region.
Human fetal brain tissue from terminations of pregnancy was provided by the Human Developmental Biology Resource, Institute of Child Health, University College London. Genomic DNA from 40 first-trimester and 100 second-trimester fetal brain samples was initially genotyped to identify heterozygotes for the ZNF804A-expressed SNP rs12476147, suitable for the allelic expression assay. Assayed first-trimester samples (heterozygous for rs12476147) were derived from the whole brain of 16 fetuses aged between 8 and 12 weeks postconception. Assayed second-trimester samples (heterozygous for rs12476147) were derived from the whole brain of 35 fetuses aged between 14 and 20 weeks postconception. There was no significant difference in age between fetal samples derived from rs1344706 heterozygotes and homozygotes. Ethical approval for the collection and distribution of this material for research was obtained from the Institute of Human Genetics, Newcastle University, and the Institute of Child Health, University College London.
Genomic DNA and total RNA were extracted from all samples using phenol/chloroform and TRI-Reagent solution (Ambion, Life Technologies, Grand Island, N.Y.), respectively. RNA samples were treated with TURBO DNase (Ambion) prior to reverse transcription and did not yield a polymerase chain reaction (PCR) product in the absence of a reverse transcription step. Two reverse transcription reactions were performed for each sample using 1 μg total RNA, random decamers (Ambion), and SuperScript III (Invitrogen, Carlsbad, Calif.). All resulting cDNA samples were diluted 1/7 prior to assay of allelic expression.
Genotyping
Genomic DNA samples were genotyped for the schizophrenia risk SNP rs1344706 and the expressed SNP rs12476147 using SNaPshot primer extension (Applied Biosystems, Foster City, Calif.).
Linkage Disequilibrium and Haplotype Analysis
Linkage disequilibrium between the schizophrenia risk SNP rs1344706 and the expressed SNP rs12476147 (used to assay ZNF804A allelic expression) was calculated separately for the 96 postmortem adult subjects and the 40 first-trimester and 100 second-trimester fetal samples using Haploview 4.0 (23). Because the brain banks from which our samples were derived do not possess information on donor ethnicity, we tested for possible population differences in ZNF804A haplotype structure that could potentially affect our results. Haplotype frequencies predicted using Haploview were used to calculate counts of the four possible rs1344706-rs12476147 haplotypes in each of our three brain cohorts and in the nonrelated parental subjects from three populations (CEU [CEPH Utah residents with ancestry from northern and western Europe], YRI [Yoruba in Ibadan, Nigeria], and CHB [Han Chinese in Beijing, China]) that were genotyped as part of the International HapMap Project (http://hapmap.ncbi.nlm.nih.gov). Predicted haplotype counts were compared between each of the brain cohorts and the three HapMap populations using 2×4 Fisher’s exact tests (see Table S1 in the data supplement accompanying the online edition of this article). There were no significant differences in haplotype frequencies between the three brain cohorts, and none of the cohorts differed from the CEU HapMap population. In contrast, haplotype frequencies in all three brain cohorts were significantly different from those in the YRI HapMap population (p<0.005 for all comparisons).
Allelic Expression Assay
We used SNP rs12476147, located in the 3′ exon of full-length ZNF804A mRNA (NM_194250), to distinguish the RNA transcribed from the two chromosomal copies of ZNF804A in individual heterozygotes. This SNP was in strong, but not perfect, linkage disequilibrium with the schizophrenia risk SNP rs1344706 in our samples (combined sample D′=0.99; r2=0.32). This enabled us to test the effect of the rs1344706 genotype on ZNF804A allelic expression by further dividing our assayed rs12476147 heterozygotes into those that were homozygous and those that were heterozygous for rs1344706. Assayable rs12476147 heterozygotes were initially identified by genotyping genomic DNA. Sequence containing rs12476147 was then PCR-amplified from the two reverse transcription reaction products for each heterozygous sample, alongside the corresponding genomic DNA samples and two negative controls. All samples from each brain region or fetal stage were assayed concurrently. PCR primers were based on a single exon sequence, designed to produce the same amplicon from both cDNA and genomic DNA: 5′-GATTTGTCCCCAGTGCTTGT-3′ and 5′-GCCTCTGGTGGATGAAAAGA-3′. Alleles of rs12476147 were discriminated and relatively quantified using SNaPshot primer extension (using the extension primer 5′-CAGTGCTTGTCATCTTCAAC-3′) after treatment of PCR product with shrimp alkaline phosphatase (GE Healthcare, Little Chalfont, United Kingdom) and Exonuclease I (New England Biolabs, Ipswich, Mass.). Primer extension reactions were electrophoresed on an Applied Biosystems 3130 Genetic Analyzer, and peak heights of allele-specific extended primers were determined using GeneMarker (SoftGenetics, State College, Pa.). The peak height for the minor frequency A allele of rs12476147 was divided by the peak height for the major frequency T allele to calculate the allele ratio for each reaction. For each plate, the average allele ratio from all genomic DNA samples was used as a correction factor for all genomic DNA and cDNA allele ratios, since this could be assumed to reflect a perfect 1:1 ratio of the two alleles and could therefore be used to correct for any inequalities in allelic representation specific to the assay (15). This procedure was performed four times for each brain sample, giving a total of eight corrected cDNA allele ratios and four corrected genomic DNA allele ratios for each sample. Statistical tests were performed using the average corrected allele ratio for each genomic DNA and cDNA sample.
Statistical Analysis
The effect of the rs1344706 genotype on ZNF804A allelic expression was assessed by comparing average corrected cDNA allele ratios from rs1344706 heterozygotes with those from rs1344706 homozygotes. We further tested whether the risk allele of rs1344706 was associated with a net increase or decrease in ZNF804A allelic expression by comparing average corrected cDNA allele ratios from rs1344706 heterozygotes with average corrected allele ratios from genomic DNA samples. We performed t tests for all comparisons using SPSS 15.0 (SPSS, Inc., Chicago). Where differences in variance were detected between comparison groups (Levene’s test, p<0.05), we used t tests that assume unequal variance. All tests were two-tailed, and p values <0.05 were considered to be significant.
Results
Figure 1 illustrates allelic expression of ZNF804A at the expressed SNP rs12476147 in the dorsolateral prefrontal cortex, hippocampus, and substantia nigra of the adult human brain. In the dorsolateral prefrontal cortex, the average corrected cDNA allele ratio for subjects who were additionally heterozygous for the schizophrenia risk SNP rs1344706 was 1.07 (range: 0.92–1.35), compared with an average ratio of 1.10 (range: 0.92–1.36) for rs1344706 homozygotes. In the hippocampus, average corrected cDNA allele ratios were 1.12 (range: 0.93–1.25) for rs1344706 heterozygotes and 1.22 (range: 1.06–1.48) for homozygotes. In the substantia nigra, average corrected cDNA ratios were 1.03 (range: 0.85–1.13) for rs1344706 heterozygotes and 1.06 (range: 0.91–1.16) for homozygotes. Allele ratios did not significantly differ between rs1344706 heterozygotes and homozygotes in any adult brain region.
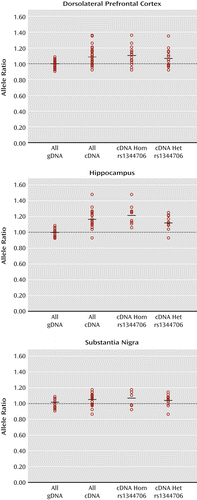
a Individual data points represent the average A/T allele ratio at the expressed single-nucleotide polymorphism (SNP) rs12476147 from four assays of genomic DNA (gDNA) or eight assays of cDNA from each heterozygous subject. All gDNA ratios, cDNA ratios, and cDNA ratios divided into rs1344706 homozygotes (Hom) and rs1344706 heterozygotes (Het) are shown. In each case, the mean allele ratio is indicated by a horizontal bar. The 1:1 allele ratio derived from the average gDNA allele ratio is indicated by a dotted line. In the dorsolateral prefrontal cortex, there is no significant difference in cDNA allele ratios between homozygotes (N=16) and heterozygotes (N=16) for the schizophrenia risk SNP rs1344706. cDNA allele ratios from both rs1344706 heterozygotes and homozygotes are significantly higher than gDNA allele ratios (p=0.002 and p=0.01, respectively, t test unequal variance). In the hippocampus, there is no significant difference in cDNA allele ratios between homozygotes (N=10) and heterozygotes (N=11) for the schizophrenia risk SNP rs1344706. cDNA allele ratios from both rs1344706 heterozygotes and homozygotes are significantly higher than gDNA allele ratios (p<0.001 for each comparison, t test unequal variance). In the substantia nigra, there is no significant difference in cDNA allele ratios between homozygotes (N=8) and heterozygotes (N=10) for the schizophrenia risk SNP rs1344706. cDNA allele ratios from both rs1344706 heterozygotes and homozygotes do not significantly differ from gDNA allele ratios.
In both the dorsolateral prefrontal cortex and hippocampus, cDNA allele ratios differed significantly from allele ratios observed in genomic DNA in rs1344706 homozygotes and heterozygotes (all p values <0.05). This suggests the presence of cis regulatory variants other than rs1344706 operating in these brain regions, which are in strong linkage disequilibrium (r2) with the assayed SNP rs12476147. Haploview analyses revealed that the risk (T) allele of rs1344706 was usually in phase with the relatively overexpressed (A) allele of rs12476147 in our adult brain samples (D′=0.92), suggesting that it is generally associated with increased ZNF804A expression in the adult brain. However, our finding that cDNA allele ratios did not significantly differ between rs1344706 heterozygotes and homozygotes in any assayed adult brain region indicates that the risk SNP does not itself substantially affect the cis regulation of ZNF804A in these regions, and therefore other cis regulatory variants are largely responsible for the observed allelic expression imbalance.
Given the apparent lack of effect of rs1344706 in the adult brain, and since schizophrenia is widely hypothesized to be a neurodevelopmental disorder (24–27), we next investigated the effect of rs1344706 on ZNF804A expression in the human fetal brain. Figure 2 illustrates allelic expression of ZNF804A at the expressed SNP rs12476147 in whole brain samples from the first and second trimester of gestation. In first-trimester brain tissue, the average corrected cDNA ratio was 1.00 (range: 0.98–1.03) for rs1344706 heterozygotes and 0.99 (range: 0.92–1.04) for homozygotes. The cDNA allele ratios did not significantly differ between the two genotype groups, nor did they significantly differ from allele ratios in genomic DNA. The small range of cDNA allele ratios that we observed strongly suggests that all common cis regulatory variants in ZNF804A are largely inactive at this developmental time point.
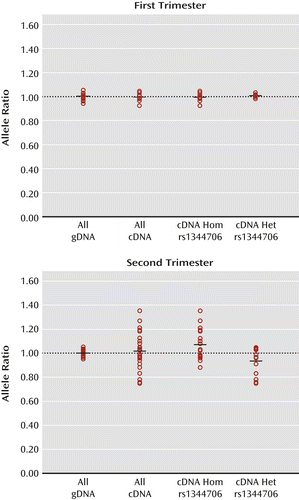
a Individual data points represent the average A/T allele ratio at the expressed single-nucleotide polymorphism (SNP) rs12476147 from four assays of genomic DNA (gDNA) or eight assays of cDNA from each heterozygous subject. All gDNA ratios, cDNA ratios, and cDNA ratios divided into rs1344706 homozygotes (Hom) and rs1344706 heterozygotes (Het) are shown. In each case, the mean allele ratio is indicated by a horizontal bar. The 1:1 allele ratio derived from the average gDNA allele ratio is indicated by a dotted line. In the first-trimester fetal brain, there is no significant difference in cDNA allele ratios between homozygotes (N=7) and heterozygotes (N=9) for the schizophrenia risk SNP rs1344706. cDNA allele ratios from both rs1344706 heterozygotes and homozygotes do not significantly differ from gDNA allele ratios. In the second-trimester fetal brain, there is a significant difference in cDNA allele ratios between homozygotes (N=21) and heterozygotes (N=14) for the schizophrenia risk SNP rs1344706 (p=0.002). cDNA allele ratios from rs1344706 homozygotes are significantly higher than gDNA allele ratios (p=0.02, t test unequal variance). cDNA allele ratios from rs1344706 heterozygotes are significantly lower than gDNA allele ratios (p=0.04, t test unequal variance). In rs1344706 heterozygotes, the relatively underexpressed A allele of rs12476147 is predicted to always be in phase (D′=1) with the schizophrenia risk (T) allele of rs1344706.
In contrast to our findings in first-trimester brain tissue, we observed a wide range of ZNF804A cDNA ratios in brain tissue from the second trimester of gestation (Figure 2). Unlike all other tested tissues, cDNA allele ratios in second-trimester brain tissue differed significantly according to rs1344706 genotype (p=0.002), averaging 0.93 (range: 0.75–1.05) in rs1344706 heterozygotes and 1.07 (range: 0.88–1.35) in homozygotes. In line with our first-trimester and adult brain samples, the risk (T) allele of rs1344706 was predicted to be in phase with the A allele of the expressed SNP rs12476147 in second-trimester rs1344706 heterozygotes (D′=1). However, unlike all other groups, in the second-trimester rs1344706 heterozygous group, the cDNA allele ratio was significantly lower than the genomic DNA allele ratio (p=0.04), indicating a generally reduced expression of ZNF804A from chromosomes carrying the risk allele of rs1344706 (compared with nonrisk). In rs1344706 homozygotes, cDNA allele ratios in second-trimester fetal brain tissue were significantly higher than genomic DNA allele ratios (p=0.02) and therefore similar to the cDNA allele ratios observed in the adult dorsolateral prefrontal cortex and hippocampus samples. Consistent with specific effects of rs1344706 genotype in fetal brain tissue, cDNA allele ratios in second-trimester rs1344706 heterozygotes were significantly different from those observed in rs1344706 heterozygotes in all assayed regions of the adult brain (p<0.05 for all comparisons).
Discussion
We assessed variable cis effects on ZNF804A expression, and the extent to which the schizophrenia risk SNP rs1344706 could account for them, in three regions of the adult human brain and in human brain tissue from two stages of fetal development. We found that although there were clear cis-acting influences on ZNF804A expression operating in the adult brain, these were not directly attributable to rs1344706. Variable cis effects on ZNF804A expression were largely absent from first-trimester brain tissue but clearly evident in samples from the second trimester of gestation. Genotype at rs1344706 was found to have a significant effect on ZNF804A allelic expression in second-trimester brain tissue, with the schizophrenia risk allele associated with reduced ZNF804A expression. Our data provide direct evidence for a molecular risk mechanism for schizophrenia operating during the second trimester of fetal brain development.
We initially tested the effect of rs1344706 on ZNF804A allelic expression in the dorsolateral prefrontal cortex, hippocampus, and substantia nigra in the adult brain. The dorsolateral prefrontal cortex and hippocampus were chosen because they have been implicated in the neurophysiological consequences of rs1344706 genotype (11, 12), and in our previous assessment of ZNF804A allelic expression in 10 adult brain regions (based on a subset of the adult subjects in the present study), the hippocampus was the region that exhibited the most pronounced allelic imbalance of ZNF804A in cDNA (21). The substantia nigra was chosen because it was the region in our previous analysis in which ZNF804A allelic expression differed most from the other regions. In the present study, we observed significant allelic expression imbalance of ZNF804A in both the hippocampus and dorsolateral prefrontal cortex, indicating heterozygous cis effects on ZNF804A expression in these two regions. However, because this was observed in rs1344706 homozygotes as well as heterozygotes, we interpret these findings as reflecting cis regulatory variants other than rs1344706 that are in strong linkage disequilibrium (r2) with the expressed SNP rs12476147.
Our findings in the adult brain are consistent with those of Williams et al. (8), who tested the effect of rs1344706 on ZNF804A allelic expression in the cerebral cortex. They reported relative overexpression of a ZNF804A allele (rs4667001-G) that is predicted to be in perfect linkage disequilibrium with the allele of rs12476147 that we found to be overexpressed in the present study (r2=1 in the CEU HapMap sample). Also consistent with our findings, Williams et al. reported no effect of rs1344706 genotype on ZNF804A allelic expression, leading them to conclude that the observed allelic expression imbalance was not directly attributable to rs1344706. Because the risk allele of rs1344706 is generally in phase with the rs12476147 and rs4667001 alleles that are overexpressed, this risk allele may be indirectly associated with increased ZNF804A expression in the adult human brain. This could explain a previously reported association between the risk allele of rs1344706 and higher total ZNF804A expression in the adult dorsolateral prefrontal cortex (5). However, these cis effects on ZNF804A expression do not appear to have a substantial effect on schizophrenia risk, since rs12476147 and rs4667001 (with which these effects are more strongly associated) are themselves only weakly associated with schizophrenia in the Williams et al. study (p=0.01) (8).
The adult brain region in which we observed the largest differences in allelic expression between rs1344706 genotypes was the hippocampus, in which we observed reduced expression of the rs12476147 allele in phase with the schizophrenia risk allele in rs1344706 heterozygotes, although the difference fell short of statistical significance (p=0.06). However, because cDNA allele ratios remained significantly higher than the (1:1) genomic allele ratios for these heterozygote subjects, it appears that any relative effect of carrying the risk allele (compared with the nonrisk allele) is generally insufficient to counter the effects of the unknown regulatory variants that increase ZNF804A expression on that haplotype.
Our finding that cDNA allele ratios in second-trimester rs1344706 homozygotes were also significantly higher than the (1:1) genomic allele ratios suggests that the (unknown) cis regulatory variants that are active in the adult brain are also active at this developmental time point. However, because cDNA allele ratios in second-trimester rs1344706 heterozygotes were significantly lower than the (1:1) genomic ratios, the rs1344706 risk allele can be seen to generally override the effects of these variants at this developmental stage, bringing about a net reduction in ZNF804A expression from chromosomes on which it is present.
Our analysis of ZNF804A haplotype frequencies (see Table S1 in the online data supplement) indicates that the observed differences between adult and fetal brain tissue samples in the effects of rs1344706 genotype are unlikely to be explained by differences in ethnicity. However, given the dynamic nature of cis effects on gene expression, we cannot exclude effects of rs1344706 at developmental stages and in tissue that we have not examined. It therefore remains possible that the effects of rs1344706 persist into later stages of fetal and postnatal life.
It should be noted that the variable cis effects on ZNF804A expression that we observed are small and would require far larger sample sizes to be reliably detected using more conventional measures of ZNF804A expression between subjects. Unlike measures of a gene’s total expression (for example, as obtained by quantitative PCR), the allelic expression method is internally controlled for trans-acting influences that can confound comparisons between subjects of different genotype because these will generally affect expression of each allele to the same extent. Potential trans-acting confounders include effects of other gene loci, environmental exposures such as drugs, and postmortem variables such as tissue or RNA quality. Importantly, measures of relative allelic expression will be influenced by trans-acting variation only when this specifically interacts with cis regulatory variation (e.g., the presence or absence of a transcription factor that binds to a variable cis regulatory site), potentially leading to allelic expression imbalance that is tissue- or temporal-specific.
The simplest explanation for our findings is that a trans regulatory factor that binds to rs1344706 makes a greater (and perhaps specific) contribution to the regulation of ZNF804A expression during fetal brain development. We previously reported that the risk allele of rs1344706 binds significantly less of an unidentified nuclear protein that is expressed in SH-SY5Y cells and in neural cell lines derived from human fetal brain (13). Data from the Human Brain Transcriptome database (http://hbatlas.org) (28, 29) reveal that ZNF804A expression peaks during the second trimester of gestation in the neocortex, striatum, hippocampus, mediodorsal thalamus, amygdala, and cerebellar cortex (see Figure S1 in the online data supplement). Data from the BrainCloud database (http://braincloud.jhmi.edu) (30) reveal a similar peak of ZNF804A expression during the second trimester of gestation in the prefrontal cortex (see Figure S2 in the online data supplement). Given that the effects of rs1344706 appear to coincide with this peak of ZNF804A expression, it could be speculated that the trans regulator that binds to sequence containing rs1344706 serves to enhance ZNF804A expression at this developmental stage.
In demonstrating a functional effect of a well-supported schizophrenia susceptibility variant in fetal brain, our data provide direct support for the hypothesis that schizophrenia is, at least in part, a neurodevelopmental disorder (24–27). The ZNF804A gene is predicted to encode a zinc finger protein, suggesting a role in the regulation of gene expression through DNA or RNA binding. We recently reported that experimental knockdown of ZNF804A in neural cells derived from human fetal cortex results in altered expression of genes involved in cell adhesion (31). The fetal down-regulation of ZNF804A expression associated with the risk allele of rs1344706 may therefore affect developmental processes that involve cell adhesion, such as neural migration, neurite outgrowth, and synaptogenesis.
Previous studies have reported associations between putative schizophrenia risk variation and the expression, in the adult brain, of gene transcripts that exhibit prominent expression in the fetal brain (e.g., 32, 33). However, the present study, to our knowledge, is the first to directly demonstrate association between a schizophrenia risk variant and altered gene expression in the fetal brain. Although we cannot rule out effects of rs1344706 at later stages of development, our data provide evidence for a risk mechanism for schizophrenia that operates long before the overt manifestation of the illness.
1 : Genetics of psychosis; insights from views across the genome. Hum Genet 2009; 126:3–12Crossref, Medline, Google Scholar
2 : Schizophrenia genetics: where next? Schizophr Bull 2011; 37:456–463Crossref, Medline, Google Scholar
3 : Identification of loci associated with schizophrenia by genome-wide association and follow-up. Nat Genet 2008; 40:1053–1055Crossref, Medline, Google Scholar
4 : Genome-wide association study identifies five new schizophrenia loci. Nat Genet 2011; 43:969–976Crossref, Medline, Google Scholar
5 : Replication of association between schizophrenia and ZNF804A in the Irish Case-Control Study of Schizophrenia sample. Mol Psychiatry 2010; 15:29–37Crossref, Medline, Google Scholar
6 : Expanding the range of ZNF804A variants conferring risk of psychosis. Mol Psychiatry 2011; 16:59–66Crossref, Medline, Google Scholar
7 : Population-based and family-based association studies of ZNF804A locus and schizophrenia. Mol Psychiatry 2011; 16:360–361Crossref, Medline, Google Scholar
8 : Fine mapping of ZNF804A and genome-wide significant evidence for its involvement in schizophrenia and bipolar disorder. Mol Psychiatry 2011; 16:429–441Crossref, Medline, Google Scholar
9 : Allelic differences between Han Chinese and Europeans for functional variants in ZNF804A and their association with schizophrenia. Am J Psychiatry 2011; 168:1318–1325Link, Google Scholar
10 : Evidence of sex-modulated association of ZNF804A with schizophrenia. Biol Psychiatry 2011; 69:914–917Crossref, Medline, Google Scholar
11 : Neural mechanisms of a genome-wide supported psychosis variant. Science 2009; 324:605Crossref, Medline, Google Scholar
12 : Altered cortical network dynamics: a potential intermediate phenotype for schizophrenia and association with ZNF804A. Arch Gen Psychiatry 2011; 68:1207–1217Crossref, Medline, Google Scholar
13 : Allelic differences in nuclear protein binding at a genome-wide significant risk variant for schizophrenia in ZNF804A. Mol Psychiatry 2011; 16:787–789Crossref, Medline, Google Scholar
14 : Allelic variation in human gene expression. Science 2002; 297:1143Crossref, Medline, Google Scholar
15 : Cis-acting variation in the expression of a high proportion of genes in human brain. Hum Genet 2003; 113:149–153Medline, Google Scholar
16 : A haplotype implicated in schizophrenia susceptibility is associated with reduced COMT expression in human brain. Am J Hum Genet 2003; 73:152–161Crossref, Medline, Google Scholar
17 : Haplotypes at the dystrobrevin binding protein 1 (DTNBP1) gene locus mediate risk for schizophrenia through reduced DTNBP1 expression. Hum Mol Genet 2005; 14:1947–1954Crossref, Medline, Google Scholar
18 : Detection of regulatory variation in mouse genes. Nat Genet 2002; 32:432–437Crossref, Medline, Google Scholar
19 : Digital RNA allelotyping reveals tissue-specific and allele-specific gene expression in human. Nat Methods 2009; 6:613–618Crossref, Medline, Google Scholar
20 : Dynamic variation in allele-specific gene expression of Paraoxonase-1 in murine and human tissues. Hum Mol Genet 2008; 17:3263–3270Crossref, Medline, Google Scholar
21 : Effects of cis-regulatory variation differ across regions of the adult human brain. Hum Mol Genet 2010; 19:4490–4496Crossref, Medline, Google Scholar
22 : Context-specific functional effects of IFNGR1 promoter polymorphism. Hum Mol Genet 2006; 15:1475–1481Crossref, Medline, Google Scholar
23 : Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 2005; 21:263–265Crossref, Medline, Google Scholar
24 : From neuropathology to neurodevelopment. Lancet 1995; 346:552–557Crossref, Medline, Google Scholar
25 : Schizophrenia: a disorder of neurodevelopment? Curr Opin Neurobiol 1997; 7:285–289Crossref, Medline, Google Scholar
26 : Schizophrenia as a disorder of neurodevelopment. Annu Rev Neurosci 2002; 25:409–432Crossref, Medline, Google Scholar
27 : Is schizophrenia a neurodevelopmental disorder? Br Med J 1987; 295:681–682Crossref, Medline, Google Scholar
28 : Functional and evolutionary insights into human brain development through global transcriptome analysis. Neuron 2009; 62:494–509Crossref, Medline, Google Scholar
29 : Spatio-temporal transcriptome of the human brain. Nature 2011; 478:483–489Crossref, Medline, Google Scholar
30 : Temporal dynamics and genetic control of transcription in the human prefrontal cortex. Nature 2011; 478:519–523Crossref, Medline, Google Scholar
31 : Knockdown of the psychosis susceptibility gene ZNF804A alters expression of genes involved in cell adhesion. Hum Mol Genet 2012; 21:1018–1024Crossref, Medline, Google Scholar
32 : Neuregulin 1 transcripts are differentially expressed in schizophrenia and regulated by 5′ SNPs associated with the disease. Proc Natl Acad Sci USA 2006; 103:6747–6752Crossref, Medline, Google Scholar
33 : A primate-specific, brain isoform of KCNH2 affects cortical physiology, cognition, neuronal repolarization and risk of schizophrenia. Nat Med 2009; 15:509–518Crossref, Medline, Google Scholar

